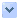我台前文<中國地效飛行船>念情曾問:發明那麼多武器幹啥/地球經得起摧殘嗎?
站長回應:人是萬物之靈嘛/地球連核子冬天(含暖化)也不會被毁/人就可能跟恐龍一樣消失了/沒了人類的地球可能會比較自然。
站長英明(真屁臭!非人的哲學家!)科學人也如是觀:
<如果地球上沒有人>科學人07/08/06【口述/魏斯曼;翻譯/陳儀蓁】
假設地球上所有的人類都消失了,世界將發生什麼變化?藉由這種創新的方式,可以審視人類如何影響環境!
經常有人想像自己是地球上最後一個倖存的人,是常有的事,不過如果所有的人類都瞬間從地球上消失了呢?美國科學作家、亞 利桑那大學新聞系副教授魏斯曼(Alan Weisman)的新書《少了我們的世界》(The World Without Us)就以此為開端。在這個思考實驗中,魏斯曼並未言明人類毀滅的原因,相反的,他直接假設人類突然從地球上消失,然後預測接下來的數年、數十年、數百 年,最有可能發生的事件。
根據魏斯曼的描述,我們大部份的基礎建設,幾乎馬上就會開始毀損,少了清道夫、道路維修人員,優美的林蔭大道、高速公路,在數個月內就會開 始出現裂痕、扭曲變形;接下來的數十年,房屋、辦公大樓陸續倒塌,但是某些一般的用品,則可能長時間都不會腐朽,例如不鋼鍋,如果埋在雜草叢生的小土丘下 (那裡曾經是我們的廚房),就可能保存數千年之久;而某些常見的塑膠,也許會保持完好長達數萬年之久,在微生物演化出消化它們的能力之前,這些塑膠是不會 分解的。
Scientific American編輯米爾斯基(Steve Mirsky)親自訪問魏斯曼,探究他寫作這本書的動機,以及我們可以從書中學到哪些教訓,以下是訪談內容的節錄。
如果明天就是人類的末日,曼哈頓宏偉壯觀的建築群在短時間內將不復存在,魏斯曼勾勒出了紐約市從水泥叢林轉變為原始森林的過程。
「如果有一天人類不在這裡了,我們所創造的那些東西會變成什麼樣呢?大自然會把我們留下的痕跡一掃而空嗎?有沒有哪些東西是無法摧毀或不可抹滅的?大自然有可能把紐約市帶回1609年、英國探險家哈德遜(Henry Hudson)當初見到的那片森林嗎?」
「我問過紐約市的工程師與維修人員,他們得花多少力氣在抵抗大自然的力量,結果十分驚人。我發現那些壯觀、氣勢凌人的建築,雖然看起來那麼 雄偉且屹立不搖,事實上卻十分脆弱,這些建築能夠存在並正常運作,多虧了少數人在支撐,而我們大多數人也都依賴著這些人,生活才得以照常運轉。『曼哈頓』 這個名字源自印第安語的『丘陵』,這裡曾是一座丘陵起伏的島嶼,當然,人們最後剷平這些丘陵,鋪上棋盤式街道。過去曾經有40條溪流貫穿這些丘陵,還有許 多泉水遍佈整座曼哈頓島,這些水現在到哪裡去了呢?這裡的雨水也跟以前一樣多,但這些水現在都受到監控,藏在地底下,有些則在污水系統中流動,但是污水系 統並不像大自然的毛細作用那樣有效,以致大部份的地下水都在地下蠢動、隨時想竄出,就算是晴朗無雲的豔陽天,地下鐵的維護人員光是一天就得抽出4920萬公升的水,否則大水就會開始淹進隧道。」
「曼哈頓有些地區得持續對抗水位不斷升高的地下河流侵蝕軌道。當你站在幫浦室裡,看著大量的水湧入,而腳下的小箱子裡,正是把水抽出的幫浦。所以說,如果 人類明天就從地球上消失,首先會發生的事情之一就是電力中斷。我們的電力主要來自核能或燃煤發電廠,這些電廠都有自動的故障安全防護開關,確保系統在無人 監控的情況下,一切不會失控。一旦電力中斷,幫浦就會停止運作,水則會灌入地下鐵。在48小時內,紐約市有很多地方會開始淹水,甚至從地面就可看到,例如 污水管會產生溢流,因為這些污水管很快就被垃圾堵住,一開始,這些堵塞物是飄散在紐約市的塑膠袋,之後,如果公園的籬笆無人修剪,就換成枯枝落葉了。」
「但是地底下會發生什麼事呢?腐蝕。想像一下你站在萊辛頓大道下方的地下鐵,等待列車進站,而上方由鋼柱支撐著的屋頂,其實就是街道。這些 鋼柱會漸漸腐蝕,最後倒塌,過一陣子街道上就會出現坑洞,這一切只要花二、三十年的時間。而且過不了多久,某些街道就會變回地表上的河流──那些在我們大 興土木之前,原本就存在於曼哈頓的河流。」
「曼哈頓有很多建築是固定在岩床上的,即使這些建築有鋼筋支撐,它們的結構也不是為浸水所設計,這些建築最終會傾斜、倒塌。而隨著氣候變 遷,天氣變得更極端,勢必有更多颶風侵襲東岸。通常一棟建築的倒塌,會連帶引起其他建築的倒塌,進而闢出一片空地。植物的種子隨風飄進這些空地,然後在路面的細縫中落腳;雖然這些種子在落葉形成的腐植層也能輕易生根,但混凝土粉碎後產生的石灰,將營造一個酸度較低的環境,適合多種的植物生存。這座城市將開始發展屬於自己的小生態系。每年春寒時節,新的裂縫就會產生,水流進這些裂縫然後結冰,導致縫細擴大,植物種子就會飄進這些新的裂縫。這些情況很快就會發生。」
一旦人類在這個世界上消失,地球的生態系會如何轉變呢?魏斯曼說,我們可以參考人類足跡罕至的那些原始地區,或許可以從中一瞥這個假想中的世界。
「為了一探人類消失後的世界會變成什麼樣子,我前往人類因為各種原因而離開的荒地,其中一處是歐洲最後一座原始森林,那裡就像小時候有人讀 格林童話給我們聽時,心中浮現的影像:一座陰森幽暗的森林,樹上掛著厚重的苔蘚,狼嚎不絕於耳。這樣的地方依然存在於波蘭和白俄羅斯的邊境,那裡是13世 紀立陶宛一位公爵設立的狩獵場,這位公爵後來成為波蘭國王,日後許多波蘭國王及俄國沙皇,均保留此地做為私人狩獵場。這座森林鮮少有人類干擾,第二次世界 大戰之後就成為國家公園。走進這座森林,舉目可見參天林木,這種感覺並不奇怪,反而令人舒暢,就像某些東西在這裡才是完整的,你可以看見45公尺高、3公 尺粗的橡樹、白蠟樹,這些樹的樹皮溝痕之深,足以讓啄木鳥塞滿松果;除了狼與麋鹿,這裡也是最後一群原生歐洲野牛的家。」
「我也到過韓國的非軍事武裝區,在這長約241公里、寬約4公里的窄長緩衝地帶,有兩支武力強大的世界級軍隊彼此對峙著。在這兩支軍隊之間的,則是無心插柳的野生動物保護區,如果不是因為這塊非軍事區,有些物種可能已經滅絕了。有時候你會聽到兩方士兵透過擴音器對彼此喊話,或是用閃光打出各自陣營的標語,而就在這樣緊張的情勢之中,你卻可以看見成群的鶴在此過冬。」
「不過,為了真正了解少了人類的世界,我知道我必須認識人類尚未演化出來的世界,所以我去了非洲,這是人類的起源地,也是唯一仍有大型動物漫步的大陸。世界各大洲及許多島嶼,都曾有大型動物的分佈,北美洲和南美洲過去有比猛象還大的大樹獺、和熊一樣大的海狸,究竟牠們為何從地球上滅絕,至今仍眾說紛紜,但矛頭大多指向人類,各大洲的滅絕事件似乎都與人類抵達的時間相符。不過在非洲,人類與動物卻共同演化,而且那裡的動物已經學會如何躲避人類的獵殺。一旦少了人類,北美洲大概會在不久的時間內成為巨鹿的棲息地,隨著森林面積逐漸復原,在演化的長河裡,大型草食動物終將演化出攝取所有木本植物養 份的能力,大型的肉食動物也將跟著演化。」……
人類演化 维基百科



人類演化(Human evolution)指的是智人這個獨特物種的起源和演化,這是相對於其他人科動物(hominid)、大猿與胎盤哺乳動物而言。對於人類演化的研究涵蓋幾個科學學科,包括體質人類學、靈長類學、考古學、語言學與遺傳學。[1]
在人類演化的脈絡下,「人類」這個專有名詞指的是「人屬」,但人類演化的研究往往包括其他人科動物,如南方猿,人屬是在大約230萬至240萬年前的非洲,從南方猿人屬分支出來。[2][3]科學家們估計,人類大約在距今500萬到700萬年之間,從他們與黑猩猩的共同祖先分支出來。其後發展的若干人屬物種和亞種,已告滅絕。這些包括棲息在亞洲的直立人、棲息在歐洲的尼安德塔人。古智人大約在距今40萬年到25萬年間演變出來。
在科學家之間,關於「解剖學上的現代人類」(anatomically modern humans)的起源,所持的主流觀點是「現代人類晚近非洲起源說」或「源出非洲」假說。[4][5][6][7]這個假說主張,智人興起於非洲,並且在大約距今5萬年到10萬年間遷移出非洲大陸,取代了在亞洲的直立人以及在歐洲的尼安德塔人。支持另一種「多區域演化」的科學家主張,智人是在地理上相區隔但維持相互混血的諸多人類群體之間所產生的,這些群體源自於在距今約250萬年前,源出非洲的直立人所從事的一場遍及世界各地的遷徙。證據顯示,尼安德塔人的數個單倍群類型出現於目前在非洲以外的人類群體中;而且,尼安德塔人及其他人屬,例如傑尼索瓦人(Denisova hominin)可能貢獻了他們 6% 的基因體給現今的人類。[8][9]
目錄
1 思想史
2 在人屬出現之前 2.1 大猿的演化
2.2 人類譜系從其他大猿分支之後
3 人屬 3.1 巧人
3.2 人屬魯道夫種與人屬喬治亞種
3.3 匠人與直立人
3.4 西布蘭諾人與人屬先驅種
3.5 人屬海德堡種
3.6 羅德西亞人與賈威斯頭骨
3.7 尼安德塔人
3.8 智人
3.9 弗洛瑞斯人
3.10 傑尼索瓦人族
4 人屬物種對照表
5 使用工具 5.1 石器
5.2 現代人類與「大躍進」論辯
6 人類演化的模型 6.1 源出非洲
6.2 多地起源模型
7 衍生閱讀
8 參考文獻
思想史
人屬的英文homo這個詞,這個用來稱呼人類所屬的生物屬名,是拉丁文稱呼「人類」的字詞。它最初是由卡爾·林奈選來用在他的分類系統中。英文的「人類」一字human源自於拉丁文的humanus,這是homo的形容詞。拉丁文的homo源自於印歐語系的語根dhghem,意指「大地」。[10]
卡爾·林奈及當時其他科學家也認為,大猿是人類最親密的近親,這是由於形態學和解剖上的相似處。人類與早期類人猿在譜繫上具有連結的這種可能性,直到1859年查爾斯·達爾文出版《物種起源》之後,才可能變得更清楚。這本書主張支持新物種由早先物種演化而來的概念。達爾文的書沒有觸及人類演化的問題,只是說:「光芒將會投射在人類起源和他的歷史」。
最早關於人類演化本質的爭辯,出現在湯瑪斯·亨利·赫胥黎和理查·歐文之間。赫胥黎認為人類從類人猿演化而來,他列舉了人類和類人猿之間的許多相似性與相異性,特別是在他1863年的書《有關人類在自然界定位的證據》(Evidence as to Man's Place in Nature)。然而,許多達爾文的早期支持者(如阿弗雷德·羅素·華萊士和查理斯·萊爾)並不同意人類心智能力和道德情感的起源,可藉由自然選擇得到解釋。達爾文於1871年發表《人類的由來》一書時,將演化理論與性選擇運用於人類本身。[11]
有一個嚴重的問題,就是缺乏介於人類與類人猿中間類型的化石。直到1920年代,這類化石才在非洲出土。1925年,雷蒙·達特(Raymond Dart)描述了南方猿人非洲種(Australopithecus africanus)的命名用標本(type specimen)是「陶格孩童」(Taung Child),這是在一座山洞裡發現的一具南方猿人嬰孩。這個孩童的遺骸包括一個保存完好的細小頭骨及其腦子的顱內腔模型(endocranial cast)。雖然這個腦子的腦容量較小(410立方公分),但是它的形狀是圓轉的,有別於黑猩猩和大猩猩的腦子,而更像是現代人類的腦子。此外,這個標本也顯示較短的犬齒,而且枕骨大孔的位置成為雙足直立行走的證據。所有這些特徵都使得達特相信,陶格孩童是一個雙足直立行走的人類祖先,也是介於類人猿與人類之間的一個過渡形式。
類人猿分類學(人類及其近親的分類)自從1950年代以來,已有很大的變化。[12]例如,纖細種南方猿人曾被認為是現代人所屬的人屬(Homo)祖先。[13]南方猿人與智人都是動物分類學上的人族(tribe Hominini)的一部分。[14]
1970年代所蒐集的資料顯示,南方猿人是個多樣化的群體,南方猿人非洲種未必是現代人類的直接祖先。[15]原先將南方猿人分類為「纖細種」與「粗壯種」,現在的重新分類將粗壯種放進單獨的屬:傍人(Paranthropus)。[15]分類學家將人類、南方猿人及相關物種,以及其他類人猿放在同一個科,稱為人科(Hominidae)。
在1980年代與1990年代DNA測序的進展,特別是粒線體DNA以及稍後的Y染色體DNA的研究,增進了對人類起緣的認識。[16][17][18]來自世界各地廣大原住民群體的粒線體DNA與 Y染色體DNA樣本排序結果,顯示了關於男性與女性基因遺傳的遠祖訊息。[19] 基因樹狀圖差異的校準,支持了現代人類晚近非洲起源說。[20] 分析呈現了再非洲各地具有最大的DNA 模式多樣性,這顯示非洲是粒線體夏娃與Y染色體亞當的祖居地。[21]
在人屬出現之前
大猿的演化
更猴

靈長類的演化史可追溯到距今6500萬年,牠們是在當時造成物種(特別是恐龍)滅絕的大災難下倖存的胎盤哺乳動物之中,最古老的一群。已知最古老的類靈長類哺乳動物物種,更猴(Plesiadapis)來自北美,但牠們遍佈於歐亞大陸和非洲,在古新世和始新世期間,這兩處地方都處於熱帶。
假熊猴

隨著當代氣候的開始,特點是在大約距今3000萬年前的漸新世第一次出現了南極冰帽。在這個時間出現的靈長類動物是假熊猴( Notharctus)。1980年代在德國發現的化石證據,確定具有1650萬年的歷史,比起來自東非的類似物種早了大約150萬年,並正在挑戰關於人類祖先源自非洲大陸的理論。
大衛·畢甘(David Begun)[22]表示,這些靈長類動物在歐亞大陸蓬勃發展,而這個譜系導向非洲猿類和人類,包括森林古猿(Dryopithecus),從歐洲或西亞向南遷移至非洲。這支後來存活下來的熱帶群體,出土最完整的一群見於開羅西南部的法揚(Fayum)堆積層,始新世晚期和漸新世最早期的化石層位之中,這引發後續所有的現生靈長類,馬達加斯加的狐猴,東南亞的懶猴,非洲的嬰猴屬(galago)或「叢林嬰兒」(bush babies),以及人形亞目(Simian anthropoids);廣鼻猴次目(platyrrhine)或新世界猴,和狹鼻猴次目(catarrhine)或舊大陸猴、類人猿和人類。
已知最早的狹鼻猴次目,來自漸新世晚期的卡莫亞猿(Kamoyapithecus),在肯亞北部大裂谷的Eragaleit出土,定年為距今2400萬年前。其祖先一般認定是來自法揚的埃及猿、原上猿與傍猿(Parapithecus),在大約距今 3500萬年前。在2010年, Saadanius 被描述為狹鼻猴次目冠狀群體(Crown group) 最後共同祖先的一個近親,目前暫時定年到距今2900萬到2800萬年間,有助於填補長達1100萬年的化石記錄斷層。[23]
在中新世早期,大約2200萬年前,來自東非的多種適應樹居生活的原始狹鼻猴亞目,顯示一段歷史悠久的早期多樣化發展。在2000萬年前的化石,包括歸類於最早的舊世界猴維多利亞猿(Victoriapithecus)的碎片。在這個屬當中,被認為處在猿類譜系之內,可推至距今1300萬年前的物種包括原康修爾猿、Rangwapithecus、 Dendropithecus、Limnopithecus、Nacholapithecus、Equatorius、Nyanzapithecus、非洲古猿 、日猿(Heliopithecus)和肯尼亞古猿,都來自東非。其他中新世的普遍非cercopithecids來自遙遠的遺址,沉積在納米比亞洞穴的Otavipithecus,出自法國、西班牙和奧地利的加泰羅尼亞皮爾勞爾猿和森林古猿,證實在中新世早中期,在氣候相對溫暖的非洲和地中海盆地,有著廣大的生物型態多樣性。年代最近的中新世類人猿,山猿,出土自義大利距今900萬年的煤層。
分子證據指出,長臂猿(長臂猿科)的譜系,大約在距今1800萬至1200萬年前,從大猿的譜系分割出來;紅毛猩猩(猩猩亞科)的譜系,大約在距今1200萬年前,從大猿的譜系分割出來。目前並未出土足以清楚記載長臂猿祖先的化石,它可能源自於一個迄今未知的東南亞類人猿群體,但化石原紅毛猩猩可由來自印度的拉瑪古猿(Ramapithecus)和來自土耳其的Griphopithecus做為其祖先的代表,追溯至距今1000萬年前。
人類譜系從其他大猿分支之後

一個雌性的阿法南方古猿(南方猿人阿法種)的重建模型。綽號「露西」。
最接近大猩猩、黑猩猩和人類最後的共同祖先,可由肯亞出土的仲山納卡里猿和希臘出土的歐蘭猿為代表。分子證據顯示,在距今800萬至400萬年前,首先是大猩猩,然後是黑猩猩,從這條通往人類的演化路線分離出來;人類的DNA大約有98.4%與黑猩猩相同,這是比較單核苷酸多態性(single nucleotide polymorphisms)所得到的結果。大猩猩和黑猩猩的化石記錄相當有限。兩者的保存狀況都不理想(熱帶雨林土壤往往是酸性,會分解骨骼)。
其他人族有可能適應赤道帶以外的乾燥環境,還有羚羊、土狼、狗、豬、大象以及馬。約在距今800萬年前,赤道帶縮小。這些化石人族 – 從與黑猩猩分道揚鑣之後的人類譜系 - 比較廣為人知。最早的是查德沙赫人(距今700萬年)和圖根原人(距今600萬年),其次是:
地猿(距今550萬到440萬年),包括地猿始祖種(Ardipithecus kadabba)和地猿根源種(Ardipithecus ramidus)。
南方猿人屬(Australopithecus)(距今400萬到180萬年),包括湖畔南方古猿、阿法南方古猿、 南方古猿非洲種、羚羊河南方古猿、驚奇南方古猿,以及源泉南方古猿。
肯亞猿人屬('Kenyanthropus)(距今300萬到270萬年)。
傍人屬(距今300萬到120萬年),包括衣索比亞傍人、鮑氏傍人、粗壯種傍人(Paranthropus robustus)。
人屬(距今200萬年至今),包括巧人、人屬魯道夫種、匠人、格魯及亞人、人屬先驅種、西布蘭諾人、直立人、海德堡人、羅德西亞人、尼安德塔人、智人伊達圖亞種、古智人、弗洛瑞斯人。
人屬
智人是人屬唯一的現生物種,另有某些已告滅絕的人屬物種可能是智人的祖先,有許多可能是我們的「表兄弟」,已經透過物種生成過程而分離出去(speciated),遠離我們祖先的路線。[24] 關於哪些群體應算作單獨的物種,哪些是亞種,目前尚未達成共識。在某些情況下,這是由於缺乏化石證據;在其他情況下,這是由於使用細微的差異,來分類人屬的各種不同物種的結果。「撒哈拉泵浦理論」(Sahara pump theory) [25](描述一個偶爾勉強稱得上「濕潤」的撒哈拉沙漠)提供對早期人屬變異的解釋。
依據考古學和古生物的證據,有可能在某種程度上推斷出,各種古代人屬的飲食習慣,並研究在人屬之中,飲食對於體質與行為演化所扮演的角色。[26][27][28][29][30]
巧人

巧人( H. habilis)存活在大約距今240萬到140萬年前。巧人是人屬的第一個物種,在 上新世晚期或 更新世早期的非洲南部與東非,大約距今250萬到200萬年前,從南方猿人分支出來。巧人具有比南方猿人更小的大臼齒與更大的腦子,並且運用石頭(可能也用動物骨骼)製作工具。這是目前已知最早的人屬動物之一,其發現者路易·李基(Louis Leakey),因著牠與石器的關連性,暱稱為「巧人」。某些科學家提議將這個物種的屬別,從人屬移到南方猿人屬,因為依據對於牠的骨架所做的型態學分析,牠比較適應於樹居(arboreal,棲息在樹上),而不是如同智人一般雙足直立行走(bipedalism)。[31]
人屬魯道夫種與人屬喬治亞種
這些物種名稱是用在大約從距今190到160萬年間的化石,然而牠們與巧人的關係尚未釐清。
人屬魯道夫種(Homo rudolfensis)指稱來自肯亞的一個不完整的頭骨。科學家建議這是另一個巧人,但這尚未獲得確認。[32]
人屬喬治亞種(Homo georgicus)來自喬治亞共和國,可能是介於巧人與直立人間的一個中間型態物種,[33]或是直立人的一個亞種。[34]
匠人與直立人
關於人科群體基於化石證據的時空分布模擬,[35]其他詮釋方式的主要差別在於人科物種的分類學與地理分布。
第一個直立人化石是由荷蘭軍醫杜保斯(Eugene Dubois)在1891年於印度尼西亞的爪哇島發現的。他起初將這批化石材料命名為Pithecanthropus erectus,這是依據這些化石的型態學,他認為這是介於人類與大猿的中間物種。[36]直立人(Homo erectus)存活的期間大約從距今180萬年到7萬年前(這或許指出,牠們可能受到距今約7萬年前的多峇巨災(Toba catastrophe)影響而滅絕;然而,直立人梭羅亞種(Homo erectus soloensis)與弗洛瑞斯人(Homo floresiensis)逃過這場巨災而存活下來。在前半段,從距今180萬到125萬年前的直立人,被認為是一個獨立的物種:匠人(Homo ergaster),或是直立人的亞種:直立人匠人亞種(Homo erectus ergaster)。
在更新世早期,距今150萬到100萬年間的非洲、亞洲與歐洲,某些巧人群體發展出較大的腦子,並製作更進步的石器;連同其他差異,足以讓人類學家將牠們分類為一個新物種:直立人。此外,直立人確實是第一個直立行走的人類祖先。[37] 這可能藉由膝蓋的演化,以及枕骨大孔(foramen magnum,人類頭骨正下方的大孔,連接腦子與脊髓)的位置前移而成為可能。直立人用火煮食肉類。
直立人的有名標本是北京人(因其出土化石數量龐大且層位較多);其他標本在亞洲(特別在印度尼西亞)、非洲與歐洲出土。現在有許多古人類學家用「匠人」來指稱這個群體的非亞洲型態,「直立人」這個名稱只保留用在亞洲所發現的化石,以呼應牠們在骨架與齒式方面有別於「匠人」之處。
西布蘭諾人與人屬先驅種
這些物種可能是介於直立人與人屬海德堡種之間的中介物種。
人屬先驅種(Homo antecessor)已知是來自西班牙與英國的化石,定年為距今120萬年到50萬年前。[38][39]
西布蘭諾人(Homo cepranensis)指稱一個在義大利出土的頭蓋骨,據估計約在距今80萬年前。[40]
人屬海德堡種
人屬海德堡種(H. heidelbergensis)存活在大約80萬到30萬年前。學者也提議將其分類為智人海德堡亞種(Homo sapiens heidelbergensis)或智人古匈牙利亞種(Homo sapiens paleohungaricus)。[41]
羅德西亞人與賈威斯頭骨
羅德西亞人(H. rhodesiensis),估計其年代約在距今30萬年到125,000年前。 目前大多數的專家相信羅德西亞人是屬於人屬海德堡種,即使其他學者提議其為古智人與智人羅德西亞亞種(Homo sapiens rhodesiensis)。
在2006年2月,賈威斯頭骨(Gawis cranium)的化石出土,這可能是介於直立人與智人之間的中間物種,或是許多已告滅絕的演化路線之一。這個來自衣索比亞賈威斯的頭骨的年代約在距今50萬年到25萬年間。目前只有出版摘要資料,尚未由這個團隊發表經過同僚審查的研究報告。賈威斯頭骨的臉部特徵顯示,牠是一個中間物種或是「波多人」(Bodo man)女性的標本。[42]
尼安德塔人

尼安德塔人(H. neanderthalensis)存活的時間從距今40萬年前。[43] 也有學者提議將其分類為智人尼安德塔亞種(Homo sapiens neanderthalensis):學者持續爭辯「尼安德塔人」是否為一個單獨物種:人屬尼安德塔種,或者是智人的一個亞種。[44] 雖然這個爭論尚未平息,來自粒線體DNA 排序的證據指出,在尼安德塔人與智人之間並未發生明顯的基因流佈。因此,這兩者是分別的物種,其共同祖先約在距今66萬年前。[45][46] 在1997年,馬克史東慶(Mark Stoneking)說道:「[依據從尼安德塔人骨骼所萃取的粒線體DNA] 得到的結果指出,尼安德塔人並未貢獻粒線體DNA給現代人類… 尼安德塔人並不是我們的祖先」。對於第二組尼安德塔人來源的調查,支持了這些發現。[47]然而,在2010對於尼安德塔人基因體的排序指出,尼安德塔人確實在大約公元前75,000年與智人混血 (在智人遷移離開非洲之後,但在他們分別遷往歐洲、中東與亞洲之前)。[48]當代人類的DNA約有1%到4%源自尼安德塔人的DNA。[48] 這個1-4% 的DNA差異只出現在非洲以外的人類。[48] 然而,「多區域演化」假說的支持者指出,最近研究顯示,非洲以外的核心DNA遺傳的時間可回溯到距今100萬年,[49]即使這些研究的可靠性受到質疑。 [50]來自智人的競爭,可能導致尼安德塔人的滅絕。[51][52]他們有可能在歐洲並存,直到距今1萬年前。[53]
智人
智人(Homo sapiens) (種名sapiens的拉丁文原意為「聰明」或「有知識」)存活於距今25萬年到現在。在大約距今40萬年至25萬年間,當時是更新世第二間冰期,人類的腦容量變大,且發展出更細緻的石器製作技術,這些都是由直立人轉變為智人的證據。直接的證據顯示,有一股直立人從非洲向外遷移,接著在非洲的物種生成過程中,智人從直立人演化出來。智人後續在非洲內部與向非洲以外地區的遷移,最終取代了較早散居各地的直立人。這個遷移與起源理論往往稱為單地起源說或非洲起源說(Out of Africa theory)。目前的證據並未排除某些多區域演化(multiregional evolution)或遷居的智人與各地既有的人屬群體混血的可能。這是目前古人類學的一個熱烈爭辯的領域。
當前的研究確認了,人類在遺傳學上是高度同質的;換言之,人類個體的DNA比起其他物種而言,具有較多的相似性。這可能源自人類演化的時間相對晚近,或由某些毀滅性的自然災害所引起的種群瓶頸,例如多峇巨災。[54][55][56]然而,人類各群體已產生獨特的遺傳特徵,主要導因於人類的小群體遷移到許多新環境。這些因適應而產生的特徵,僅佔智人基因體的極小成分,但包括了各種不同特徵,例如膚色與鼻型,此外也有內部構造的特色,例如在高海拔地區更有效呼吸的能力。
來自衣索比亞的智人伊達圖亞種可能是一個已消失的亞種,存活於大約距今16萬年前。
弗洛瑞斯人

弗洛瑞斯人(H. floresiensis)大約存活在距今10萬年到12,000年間,由於體型嬌小,被暱稱為哈比人,這可能是島內侏儒化所造成的結果。[57] 弗洛瑞斯人引人好奇的是牠們的體型與年代,這是一個具體的晚近的人屬物種,但展現了許多當代人類所沒有的衍生特徵(derived traits)。換言之,弗洛瑞斯人與當代人類具有共同祖先,但從當代人類的譜系分出去,依循一條獨特的演化路徑。主要的發現是一個年約30歲的女性骨架。在2003年出土,定年約在距今18,000年。這名女子存活時身高約一公尺,腦容量約380立方公分。(其腦容量如同黑猩猩,不到智人的平均值1400立方公分的三分之一。)
然而,學界持續爭辯著,究竟「弗洛瑞斯人」是否確實是個單獨存在的物種。[58] 目前有一些科學家相信,「弗洛瑞斯人」是受到病理侏儒症所苦的一種「智人」。[59]這一假說受到部分的支持,因為在弗洛瑞斯島,也就是這個骨架被發現的島嶼,有某些當代人類是俾格米人。有人認為,這些較矮的俾格米人再加上病理侏儒症,確實可能創造出一種類似哈比人的人類。其他主要對於「弗洛瑞斯人」這個物種概念所做的攻擊,在於在其出土地點的工具僅僅與智人有關。[59]
但是,這個病理侏儒症的假說無法解釋的是,為何弗洛瑞斯人其他的解剖特徵並不像現代人類(無論患病與否),反而很像我們種屬的古代成員。除了頭骨特徵之外,這些特徵也包括手腕、前臂、肩膀、膝蓋和腳的骨骼形式。
傑尼索瓦人族
2008年,在西伯利亞阿爾泰山從事研究工作的考古學家,於傑尼索瓦洞穴發現一個小指骨頭碎片,來自目前稱為傑尼索瓦人族(Denisova hominin)群體的一位青少年。[60]在這處洞穴的相同層位出土的器物,包括一個手鐲,依據放射性碳定年法大約在距今四萬年前。由於傑尼索瓦洞穴氣候涼爽,因此在化石片段之中的DNA得以保存下來,目前已對其粒線體DNA及核心基因組DNA完成排序。[61][8]
雖然其粒線體DNA的分歧點時間深度遠比預期更深,[62]但是完整的基因組排序,顯示傑尼索瓦人與尼安德塔人屬於相同的世系,但兩者就在現代人類興起不久之後,就從同一條路線分裂開來。[8]目前已知在歐洲,現代人類與尼安德塔人的重疊時間超過一萬年,而且這項發現引發一種可能性,尼安德塔人、現代人類、傑尼索瓦人或許曾經並存。帕博(Pääbo)指出,這個分支的存在,使得更新世晚期的圖像更加複雜。[60]
人屬物種對照表人屬物種對照表|
| 人屬先驅種 Homo antecessor | 1.2 – 0.8 | 西班牙 | 1.75公尺(6英尺) | 90公斤(198磅) | 1,000 | 2處遺址 | 1997 |
|---|
| 西布蘭諾人Homo cepranensis | 0.9 – 0.8? | 義大利 | | | 1,000 | 1 個頭蓋骨 | 1994/2003 |
|---|
| 直立人Homo erectus | 1.5 – 0.2 | 非洲, 歐亞大陸 (爪哇, 中國, 印度, 高加索) | 1.8公尺(6英尺) | 60公斤(132磅) | 850 (早期) – 1,100 (晚期) | 多處遺址 | 1891/1892 |
|---|
| 匠人Homo ergaster | 1.9 – 1.4 | 東非與南非 | 1.9公尺(6英尺) | | 700–850 | 多處遺址 | 1975 |
|---|
| 弗洛瑞斯人Homo floresiensis | 0.10? – 0.012 | 印尼 | 1.0公尺(3英尺) | 25公斤(55磅) | 400 | 7具個體 | 2003/2004 |
|---|
| Homo gautengensis | >2 – 0.6 | 南非 | 1.0公尺(3英尺) | | | | 2010 |
|---|
| 人屬喬治亞種Homo georgicus | 1.8 | 喬治亞(國家) | | | 600 | 4具個體 | 1999/2002 |
|---|
| 巧人Homo habilis | 2.3 – 1.4 | 非洲 | 1.0–1.5 米 | 33–55 公斤 | 510–660 | 多處遺址 | 1960/1964 |
|---|
| 人屬海德堡種Homo heidelbergensis | 0.6 – 0.35 | 歐洲, 非洲, 中國 | 1.8公尺(6英尺) | 60公斤(132磅) | 1,100–1,400 | 多處遺址 | 1908 |
|---|
| 尼安德塔人Neanderthal | 0.35 – 0.03 | 歐洲, 西亞 | 1.6公尺(5英尺) | 55–70公斤 | 1,200–1,900 | 多處遺址 | (1829)/1864 |
|---|
| 人屬羅德斯種Homo rhodesiensis | 0.3 – 0.12 | 尚比亞 | | | 1,300 | 非常少 | 1921 |
|---|
| 人屬魯道夫種Homo rudolfensis | 1.9 | 肯亞 | | | | 1個頭骨 | 1972/1986 |
|---|
| 智人伊達圖亞種Homo sapiens idaltu | 0.16 – 0.15 | 衣索比亞 | | | 1,450 | 3 個頭蓋骨 | 1997/2003 |
|---|
| 現代人類Homo sapiens sapiens | 0.2 – 現代 | 世界各地 | 1.4米–1.9米 | 50–100公斤 | 1,000–1,850 | 目前仍存活 | —/1758 |
|---|
使用工具

「尖石」,這是一個奧度瓦伊礫石器,人類石器的最基本類型。
火是人類最偉大的發現之一,對於人類演化非常重要。
從英國肯特郡出土的Acheulean 手斧,這是直立人使用的燧石器。
維倫多爾夫的維納斯,舊石器時代藝術的範例。
使用工具已被學者詮釋成一種智力的標誌,而且學者提出理論,工具使用可能觸發了人類演化的某些層面----最值得一提的是人類腦子的持續擴張。古生物學目前依然未能解釋腦子經過數百萬年的擴張,即使說從能量消耗來看,腦子的需求非常大。當代人類的腦子消耗約 20瓦(每天400千卡),佔人體能量消耗的五分之一。工具使用的增加,使得人類能獵得能量豐富的肉類,並有能力處理更多能量豐富的植物產品。研究人員認為,早期人類是在演化壓力下,提高他們創造和使用工具的能力。[63]
值得一提的是,有許多物種能夠製作和使用工具,但人類這個物種控制了製作的範圍並使用更複雜的工具。目前已知最古老的工具來自衣索比亞的「奧度瓦伊石器」(Oldowan Stone Tools,發現地點為奧度瓦伊 Olduwai)。這些工具的年代距今250萬年到260萬年,來自已知最早的「人屬」物種。目前尚無證據顯示,距今250萬年前有任何「人屬」標本。在一些奧度瓦伊工具附近,出土了一個人屬化石,值得注意的是其定年在距今230萬年,這隱約顯示,也許早期人屬物種確實創造和使用這些工具。這確實可能,但沒有堅實的證據。伯納伍德(Bernard Wood)指出,「傍人屬」與這個「奧度瓦伊技術叢」("Oldowan Industrial Complex")大致同時存在這個區域。雖然沒有直接證據可佐證傍人屬是工具製造者,但他們的解剖構造提供了間接證據,顯示他們具備這方面的能力。大多數古人類學家都同意,確實最有可能是早期的「人屬」物種製作出奧度瓦伊工具。他們認為,當大多數的奧度瓦伊工具與人類化石同時被發目前,出現的始終是人屬,但不是傍人屬。[64]
1994年,蘭德爾·蘇斯曼採用人屬及傍人屬的對向性拇指來做為他的立論基礎,推論他們都是工具製造者。他比較了人類和黑猩猩的拇指骨骼和肌肉,發現人類具有三條黑猩猩所欠缺的肌肉。人類也有較厚實的掌骨與更大的頭部,這使得人類的手在精準抓握方面,比黑猩猩更成功。蘇斯曼提出辯護,人類拇指的現代解剖學構造是基於製作與操作工具的需求而產生的演化回應,這兩個物種確實是工具製造者。[64]
石器

從英國肯特郡出土的Acheulean 手斧,這是直立人使用的燧石器。經證實,最早的石器約在距今260萬年前,當時在東非的巧人使用的工具,是由圓形礫石簡單捶打而成的礫石器、砍器。[65]這標示了舊石器時代的開始,結束在最後一次冰河時代告終之時,大約距今一萬年前。舊石器時代細分為舊石器時代早期(約在距今35萬至30萬年前結束),舊石器時代中期(直到距今五萬年到三萬年前)和舊石器時代晚期。
從距今70萬至30年的這段時期,稱為阿歇爾傳統〈Acheulean〉,當時的匠人(或直立人)由燧石和石英石製作手斧,起初相當粗糙(早期阿歇爾),後來經過在石片兩側更多細部修整。在距今35萬年之後,更細緻的勒伐技術(Levallois)發展出來。它包含一系列連續敲擊,製作出刮削器、切割器、針、扁針。[65]最後,大約在距今五萬年之後,尼安德塔人及克羅馬儂人製作出更細膩與專門化的燧石工具(刀、石瓣器)。在此期間,他們開始製作骨製工具。
現代人類與「大躍進」論辯
直到大約距今五萬年到四萬年前,石器的使用似乎有一步一步的發展。每個階段(巧人、匠人、尼安德塔人)一開始都比前一個階段具有更高的水準,但一旦這個階段展開之後,進一步的發展就變得遲緩。這些「人屬」物種在文化上是保守的,但在距今五萬年之後,現代人類文化改變的速率開始增加。《第三種黑猩猩》的作者賈德·戴蒙和一些人類學家稱這個現象為「行為現代性」或「大躍進」。
現代人類開始埋葬死者、製作衣服、發展複雜的狩獵技巧(如使用誘捕坑或逼使動物跳下峭壁),並從事石洞壁畫。[66] 隨著人類文化的進展,人類的不同人群推出新穎的技術,加入既有的技術當中:如魚鉤、鈕扣和骨針等器物已經顯示不同人群之間的差異,這些器物在距今五萬年前尚未出現於人類文化中。一般來說,尼安德塔人的各個群體之間並無技術上的差異。
人類學家指出的現代人類行為的具體例證,包括專業化的工具、珠寶首飾和圖像的使用(如洞穴壁畫)、生存空間的組織、儀式(例如,有大量陪葬品的墓葬)、專門的狩獵技術、探索較不宜人的地理區域、以物易物的貿易網路等等。學者之間的爭辯還在持續進行,圍繞在究竟是一場「革命」導致現代人類的產生(人類意識的大爆炸),或者說這個演變是漸進的。[67]
人類演化的模型
今天,所有的人類都同屬單一的群體:智人智人亞種(Homo sapiens sapiens),並沒有物種區隔。然而,根據「源出非洲」模型,這並不是人屬的第一個物種:人屬的第一個物種「巧人」,在至少距今兩百萬年演變出來,巧人生活在非洲不同區域的時間相對較短,「直立人」在超過距今180萬年前演變出來,到了距今150萬年前就已散布到舊世界各地。
人類學家對於當代人類群體演化有著分歧的看法,是否只在東非發生演變、物種形成,然後遷移到非洲以外,並取代歐亞大陸的人類群體(所謂的「源出非洲」模型或「完全替代」模型);或是演變成一個相互連結的群體(由「多區域演化」假設所假定的模型)。
源出非洲
粒線體DNA的分支,這種DNA只透過母親傳給女兒。[68]
依據由克里斯·斯特林格和彼得·安德魯斯所發展的源出非洲模型(Out-of-Africa model),現代智人是在距今 20 萬年前的非洲演變而來的。智人大約在距今七萬年到五萬年間,開始從非洲遷移出來,並最終取代當時存在於歐洲和亞洲的原始人屬物種。[69][70]源出非洲已獲得研究的支持,包括對女性的粒線體DNA(mtDNA)和男性 Y染色體的研究。使用 133 個類型的粒線體 DNA 構建譜系樹狀分析後,研究人員得出結論,她們都是來自非洲的同一個女人的後代,這個女人稱為粒線體夏娃。源出非洲模型也從非洲人群具有最高的粒線體遺傳多樣性這項事實得到支持。[71]
關於這個向外遷徙的過程究竟是一次或多次,學者抱持著不同的理論。一套多次遷徙模型,其中包括向南擴散理論,[72]近年來已經獲得來自基因、語言和考古證據的支持。在這個理論中,大約在距今七萬年前,現代人類從非洲之角展開一場沿著海岸的遷徙。這群人有助於將人類擴展到東南亞和大洋洲,這解釋了為何早期人類在這些區域的遺址年代,遠遠早於在黎凡特的遺址年代。人類的第二波遷徙浪潮跨越了西奈半島進入亞洲,產生了在歐亞大陸的大批人群。第二波的人群擁有更成熟的工具技術,而且比起第一波的人群更少依賴沿海的食物來源。第一波人群的擴張證據可能已被每一次冰河時期結束時,上升的海平面所摧毀。[72]這個多次遷徙模型與來自粒線體 DNA 的研究結果相衝突,後者顯示歐亞大陸的人群以及東南亞與大洋洲的人群都是從同一個粒線體 DNA 譜系傳承下來,這支持了單一的源出非洲遷徙過程,產生了所有在非洲以外的人類群體。[73]
莎拉·提胥可夫率領的廣大非洲的遺傳多樣性研究團隊,從113個不同人群的採樣當中,發現布希曼人具有最大的遺傳多樣性,這使得他們成為14個人類基因叢集的其中一個。這項研究也定位了非洲西南部現代人類遷移的源頭,接近納米比亞和安哥拉海岸。 [74]
根據人類學家和考古學家所採用的多峇巨災理論,大約在距今七萬年前,印尼蘇門答臘島的多峇湖發生一場超級火山噴發,造成了全球性的後果,[75]致使當時的大多數人死亡,並創造出一個種群瓶頸,影響現今所有人類的基因遺傳。[76]
多地起源模型
「多地起源模型」是一種「解釋人類演化的模型」,由沃波夫(Milford H. Wolpoff)於1988年提出。[77][78]多區域演化的觀點主張,人類演化是從距今250萬年前的更新世開始,直到現在,一直維持著單一的、連續的人類物種,在世界各地演化成為現代智人。 依據多區域假設,化石及基因體資料是說明全球性人類演化的證據,這與最近由晚近非洲起源說所假定的物種形成過程相矛盾。理查李基(Richard Leakey)認為化石證據並不充足,無法解決這個爭論。 [79] 對於Y染色體及粒線體DNA的單倍群研究已大大支持一個晚近的非洲起源。 [80] 來自常染色體DNA的證據也支持晚近非洲起源具有0.000 000 000 000 000 01 的可能性。[81] 然而,某些研究顯示,古智人(Archaic Homo sapiens)的遺傳成分也有可能摻雜在當代人類當中。[82] 最近關於尼安德塔人[83]與丹尼索瓦人[84] 的基因體排序也顯示確實發生某些混雜。在非洲以外的現代人類在其基因體中,有2-4%的尼安德塔人的等位基因,美拉尼西亞人額外具有4-6%的丹尼索瓦人等位基因。這些新的結果並不與《源出非洲》模型相抵觸,但顯示發生了一些額外的混雜事件。可能源自於距今73,000年前的多峇超級火山災難的一個種群瓶頸之後,經過一段復甦時間,一支很小的群體離開非洲,並遇到尼安德塔人,可能是在中東,他們的後代擴散到世界各地,有一小支遇到了丹尼索瓦人,可能是在東南亞,就在他們遷入美拉尼西亞之前。[85]
衍生閱讀
人類演化歷程
早期人類對火的使用
人類起源
人類浩大的家族史展覽
參考文獻
1.^ Heng HH. The genome-centric concept: resynthesis of evolutionary theory. Bioessays. May 2009, 31 (5): 512–25. doi:10.1002/bies.200800182. PMID 19334004.
2.^ Stringer, C.B.. Evolution of early humans//Steve Jones, Robert Martin & David Pilbeam (eds.). The Cambridge Encyclopedia of Human Evolution. Cambridge: Cambridge University Press. 1994: 242. ISBN 0-521-32370-3. Also ISBN 0-521-46786-1 (paperback)
3.^ McHenry, H.M. Human Evolution//Michael Ruse & Joseph Travis. Evolution: The First Four Billion Years. Cambridge, Massachusetts: The Belknap Press of Harvard University Press. 2009: 265. ISBN 978-0-674-03175-3.
4.^ Out of Africa Revisited - 308 (5724): 921g - Science. Sciencemag.org. 2005-05-13 [2009-11-23]. doi:10.1126/science.308.5724.921g.
5.^ Nature. Access : Human evolution: Out of Ethiopia. Nature. 2003-06-12 [2009-11-23].
6.^ Origins of Modern Humans: Multiregional or Out of Africa?. ActionBioscience [2009-11-23].
7.^ Modern Humans - Single Origin (Out of Africa) vs Multiregional. Asa3.org [2009-11-23].
8.^ 8.0 8.1 8.2 Reich D, Green RE, Kircher M, et al.. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature. December 2010, 468 (7327): 1053–60. doi:10.1038/nature09710. PMID 21179161.
9.^ Noonan JP. Neanderthal genomics and the evolution of modern humans. Genome Res.. May 2010, 20 (5): 547–53. doi:10.1101/gr.076000.108. PMID 20439435.
10.^ dhghem. The American Heritage Dictionary of the English Language ([失效連結]). 4th. Houghton Mifflin Company. 2000.
11.^ Darwin, Charles. The Descent of Man, and Selection in Relation to Sex. Princeton, New Jersey: Princeton University Press. 1871. This edition published 1981, with Introduction by John Tyler Bonner & Robert M. May. ISBN 0-691-02369-7..
12.^ Walter Carl Hartwig. The primate fossil record. Cambridge University Press. 2002: 409 [6 November 2011]. ISBN 978-0-521-66315-1.
13.^ Frederick Grine. Evolutionary History of the "Robust" Australopithecines. Transaction Publishers. 31 December 2007: 301 [6 November 2011]. ISBN 978-0-202-36137-6.
14.^ Richard Dawkins; Yan Wong. The Ancestor's Tale: A Pilgrimage to the Dawn of Evolution. Houghton Mifflin Harcourt. September 2005: 86 [6 November 2011]. ISBN 978-0-618-61916-0.
15.^ 15.0 15.1 Philip Briggs. Tanzania: With Zanzibar, Pemba & Mafia. Bradt Travel Guides. 1 October 2009: 6 [6 November 2011]. ISBN 978-1-84162-288-0.
16.^ Amade M'charek. The Human Genome Diversity Project: an ethnography of scientific practice. Cambridge University Press. 2005: 96 [10 November 2011]. ISBN 978-0-521-83222-9.
17.^ Rob DeSalle; Ian Tattersall. Human origins: what bones and genomes tell us about ourselves. Texas A&M University Press. 2008: 146 [10 November 2011]. ISBN 978-1-58544-567-7.
18.^ R. J. Trent. Molecular medicine: an introductory text. Academic Press. 2005: 6– [9 November 2011]. ISBN 978-0-12-699057-7.
19.^ Donovan Webster; Spencer Wells. Meeting the Family: One Man's Journey Through His Human Ancestry. National Geographic Books. 20 April 2010: 53 [10 November 2011]. ISBN 978-1-4262-0573-6.
20.^ Michael R. Speicher; Stylianos E. Antonarakis; Arno G. Motulsky. Vogel and Motulsky's Human Genetics: Problems and Approaches. Springer. 3 February 2010: 606 [10 November 2011]. ISBN 978-3-540-37653-8.
21.^ Dr Kutty. Adam's Gene and the Mitochondrial Eve. Xlibris Corporation. 14 September 2009: 40 [9 November 2011]. ISBN 978-1-4415-0729-7.
22.^ Kordos L, Begun DR. Primates from Rudabánya: allocation of specimens to individuals, sex and age categories. J. Hum. Evol.. 2001, 40 (1): 17–39. doi:10.1006/jhev.2000.0437. PMID 11139358.
23.^ Zalmout, I.S.; Sanders, W.J.; MacLatchy, L.M.; Gunnell, G.F.; Al-Mufarreh, Y.A.; Ali, M.A.; Nasser, A.-A.H.; Al-Masari, A.M. et al.. New Oligocene primate from Saudi Arabia and the divergence of apes and Old World Monkeys. Nature. 2010, 466 (7304): 360–364. doi:10.1038/nature09094. PMID 20631798.
24.^ Strait DS, Grine FE, Moniz MA. A reappraisal of early hominid phylogeny. J. Hum. Evol.. 1997, 32 (1): 17–82. doi:10.1006/jhev.1996.0097. PMID 9034954.
25.^ http://en.wikipedia.org/wiki/Sahara_pump_theory
26.^ Walker, Alan. Early Hominin Diets: Overview and Historical Perspectives//Ungar, Peter S.. Evolution of the Human Diet: The Known, the Unknown, and the Unknowable. US: Oxford University Press. 2006: 3–10. ISBN 0195183460. (scroll up to view chapter 1 & part of chapter 2, which is a serendipitous result from another search. Subsequent attempts get a targeted search result gave returns without chapter 1).
27.^ Ungar, Peter S. & Teaford, Mark F.. Human Diet: Its Origin and Evolution. Westport, CT: Bergin & Garvey. 2002: 206. ISBN 0897897366.
28.^ Bogin, Barry. The evolution of human nutrition//Romanucci-Ross, Lola; Moerman, Daniel E.; & Tancredi, Laurence R.. The Anthropology of Medicine: From Culture to Method. 3. South Hadley, Mass.: Bergen and Garvey. 1997: 96–142. ISBN 0897895169.
29.^ Barnicot NA. Human nutrition: evolutionary perspectives. Integr Physiol Behav Sci. 2005, April/June, 40 (2): 114–17. doi:10.1007/BF02734246. PMID 17393680.
30.^ Leonard WR, Snodgrass JJ, Robertson ML. Effects of brain evolution on human nutrition and metabolism (PDF). Annu Rev Nutr.. 2007, 27: 311–27 [2008-12-29]. doi:10.1146/annurev.nutr.27.061406.093659. PMID 17439362.
31.^ Wood, B. & Collard, M. (1999) The changing face of Genus Homo. Evol. Anth. 8(6) 195-207
32.^ Wood B. 'Homo rudolfensis' Alexeev, 1986-fact or phantom?. J. Hum. Evol.. 1999, 36 (1): 115–8. doi:10.1006/jhev.1998.0246. PMID 9924136.
33.^ Gabounia L. de Lumley M. Vekua A. Lordkipanidze D. de Lumley H.. Discovery of a new hominid at Dmanisi (Transcaucasia, Georgia). Comptes Rendus Palevol,. 2002, 1 (4): 243–53. doi:10.1016/S1631-0683(02)00032-5.
34.^ Lordkipanidze D, Vekua A, Ferring R, et al.. A fourth hominin skull from Dmanisi, Georgia. The anatomical record. Part A, Discoveries in molecular, cellular, and evolutionary biology. 2006, 288 (11): 1146–57. doi:10.1002/ar.a.20379. PMID 17031841.
35.^ Genetic Analysis of Lice Supports Direct Contact between Modern and Archaic Humans Reed DL, Smith VS, Hammond SL, Rogers AR, Clayton DH PLoS Biology Vol. 2, No. 11, e340 doi:10.1371/journal.pbio.0020340 http://biology.plosjournals.org/perlserv/?request=slideshow&type=figure&doi=10.1371/journal.pbio.0020340&id=15540
36.^ Turner W. On M. Dubois' Description of Remains recently found in Java, named by him Pithecanthropus erectus: With Remarks on so-called Transitional Forms between Apes and Man. Journal of anatomy and physiology. 1895, 29 (Pt 3): 424–45. PMID 17232143.
37.^ Spoor F, Wood B, Zonneveld F. Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion. Nature. 1994, 369 (6482): 645–8. doi:10.1038/369645a0. PMID 8208290.
38.^ Bermúdez de Castro JM, Arsuaga JL, Carbonell E, Rosas A, Martínez I, Mosquera M. A hominid from the lower Pleistocene of Atapuerca, Spain: possible ancestor to Neandertals and modern humans. Science. 1997, 276 (5317): 1392–5. doi:10.1126/science.276.5317.1392. PMID 9162001.
39.^ Carbonell, Eudald; José M. Bermúdez de Castro et al.. The first hominin of Europe. Nature. 2008-03-27, 452 (7186): 465–469 [2008-03-26]. doi:10.1038/nature06815. PMID 18368116.
40.^ Manzi G, Mallegni F, Ascenzi A. A cranium for the earliest Europeans: phylogenetic position of the hominid from Ceprano, Italy. Proc. Natl. Acad. Sci. U.S.A.. 2001, 98 (17): 10011–6. doi:10.1073/pnas.151259998. PMID 11504953.
41.^ Czarnetzki, A. Palaeopathological and variant conditions of the Homo heidelbergensis type specimen (Mauer, Germany). Journal of Human Evolution. 2003, 44: 479. doi:10.1016/S0047-2484(03)00029-0.
42.^ Indiana University. Scientists discover hominid cranium in Ethiopia, 新聞稿. March 27, 2006 [2006-11-26].
43.^ DOI:10.1111/j.1469-185X.2008.00071.x
本引用來源將會在數分鐘後自動完成。您可以插隊或手動擴充
44.^ Harvati K. The Neanderthal taxonomic position: models of intra- and inter-specific craniofacial variation. J. Hum. Evol.. 2003, 44 (1): 107–32. doi:10.1016/S0047-2484(02)00208-7. PMID 12604307.
45.^ Krings M, Stone A, Schmitz RW, Krainitzki H, Stoneking M, Pääbo S. Neandertal DNA sequences and the origin of modern humans. Cell. 1997, 90 (1): 19–30. doi:10.1016/S0092-8674(00)80310-4. PMID 9230299.
46.^ Green RE, et al.. A Complete Neandertal Mitochondrial Genome Sequence Determined by High-Throughput Sequencing. Cell. 2008, 134 (3): 416–426. doi:10.1016/j.cell.2008.06.021. PMID 18692465.
47.^ Serre D, Langaney A, Chech M, et al.. No evidence of Neandertal mtDNA contribution to early modern humans. PLoS Biol.. 2004, 2 (3): E57. doi:10.1371/journal.pbio.0020057. PMID 15024415.
48.^ 48.0 48.1 48.2 Jennifer Viegas. Neanderthals, humans interbred, DNA proves. Discovery News. 2010-05-06 [2010-08-17].
49.^ Gutiérrez G, Sánchez D, Marín A. A reanalysis of the ancient mitochondrial DNA sequences recovered from Neandertal bones. Mol. Biol. Evol.. 2002, 19 (8): 1359–66. PMID 12140248.
50.^ Hebsgaard MB, Wiuf C, Gilbert MT, Glenner H, Willerslev E. Evaluating Neanderthal genetics and phylogeny. J. Mol. Evol.. 2007, 64 (1): 50–60. doi:10.1007/s00239-006-0017-y. PMID 17146600.
51.^ Diamond, Jared. The Third Chimpanzee: The Evolution and Future of the Human Animal. Harper Perennial. 1992. ISBN 0060984031.
52.^ How Neanderthals met a grisly fate: devoured by humans. The Observer. May 17, 2009.
53.^ "DNA identifies new ancient human dubbed 'X-woman'". BBC News. March 25, 2010.
54.^ Supervolcanoes, BBC2, 3 February 2000
55.^ Stanley H. Ambrose. Late Pleistocene human population bottlenecks, volcanic winter, and differentiation of modern humans. Journal of Human Evolution. 1998, 34 (6): 623–651. doi:10.1006/jhev.1998.0219. PMID 9650103.
56.^ Ambrose, Stanley H.. Volcanic Winter, and Differentiation of Modern Humans. Bradshaw Foundation [2006-04-08].
57.^ Brown P, Sutikna T, Morwood MJ, et al.. A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature. 2004, 431 (7012): 1055–61. doi:10.1038/nature02999. PMID 15514638.
58.^ Argue D, Donlon D, Groves C, Wright R. Homo floresiensis: microcephalic, pygmoid, Australopithecus, or Homo?. J. Hum. Evol.. 2006, 51 (4): 360–74. doi:10.1016/j.jhevol.2006.04.013. PMID 16919706.
59.^ 59.0 59.1 Martin RD, Maclarnon AM, Phillips JL, Dobyns WB. Flores hominid: new species or microcephalic dwarf?. The anatomical record. Part A, Discoveries in molecular, cellular, and evolutionary biology. 2006, 288 (11): 1123–45. doi:10.1002/ar.a.20389. PMID 17031806.
60.^ 60.0 60.1 Sample, Ian. New species of human ancestor found in Siberia. The Guardian. March 24, 2010 (原始內容存檔於2010-11-22)
61.^ Krause, Johannes; Fu, Qiaomei; Good, Jeffrey M.; Viola, Bence; Shunkov, Michael V.; Derevianko, Anatoli P.; Pääbo, Svante. The complete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature. 2010, 464 (7290): 894–897. doi:10.1038/nature08976. PMID 20336068
62.^ Katsnelson, Alla. New hominin found via mtDNA. The Scientist. March 24, 2010 (原始內容存檔於2010-11-22)
63.^ Gibbons, Ann. Solving the Brain's Energy Crisis. Science. 1998, 280 (5368): 1345–47. doi:10.1126/science.280.5368.1345. PMID 9634409.
64.^ 64.0 64.1 Freeman, Scott; Jon C. Herron. Evolutionary Analysis (4th ed.)., Pearson Education, Inc. (2007). ISBN 978-0-13-227584-2 pages 786-788
65.^ 65.0 65.1 Plummer T. Flaked stones and old bones: Biological and cultural evolution at the dawn of technology. Am. J. Phys. Anthropol.. 2004, Suppl 39: 118–64. doi:10.1002/ajpa.20157. PMID 15605391.
66.^ Ambrose SH. Paleolithic technology and human evolution. Science. 2001, 291 (5509): 1748–53. doi:10.1126/science.1059487. PMID 11249821.
67.^ Mcbrearty S, Brooks AS. The revolution that wasn't: a new interpretation of the origin of modern human behavior. J. Hum. Evol.. 2000, 39 (5): 453–563. doi:10.1006/jhev.2000.0435. PMID 11102266.
68.^ Behar et al. 2008, Gonder et al. 2007, Reed and Tishkoff.
69.^ Modern Humans Came Out of Africa, "Definitive" Study Says. News.nationalgeographic.com. 2010-10-28 [2011-05-14].
70.^ Stringer CB, Andrews P. Genetic and fossil evidence for the origin of modern humans. Science. March 1988, 239 (4845): 1263–8. doi:10.1126/science.3125610. PMID 3125610.
71.^ Cann RL, Stoneking M, Wilson AC. Mitochondrial DNA and human evolution. Nature. 1987, 325 (6099): 31–6. doi:10.1038/325031a0. PMID 3025745 (原始內容存檔於2010-11-22).
72.^ 72.0 72.1 Searching for traces of the Southern Dispersal, by Dr. Marta Mirazón Lahr, et al.
73.^ Macaulay, V.; Hill, C; Achilli, A; Rengo, C; Clarke, D; Meehan, W; Blackburn, J; Semino, O et al.. Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes. Science. 2005, 308 (5724): 1034–6. doi:10.1126/science.1109792. PMID 15890885.
74.^ Gill, Victoria. Africa's genetic secrets unlocked. BBC News. May 1, 2009 [June 8, 2011]. the results were published in the online edition of the journal Science.
75.^ "The new batch - 150,000 years ago". BBC - Science & Nature - The evolution of man.
76.^ When humans faced extinction. BBC. 2003-06-09 [2007-01-05] (原始內容存檔於2010-11-22).
77.^ Wolpoff, MH; Hawks J, Caspari R. Multiregional, not multiple origins. Am J Phys Anthropol. 2000, 112 (1): 129–36. doi:10.1002/(SICI)1096-8644(200005)112:1<129::AID-AJPA11>3.0.CO;2-K. PMID 10766948.
78.^ Wolpoff, MH; JN Spuhler, FH Smith, J Radovcic, G Pope, DW Frayer, R Eckhardt, and G Clark. Modern Human Origins. Science. 1988, 241 (4867): 772–4. doi:10.1126/science.3136545. PMID 3136545.
79.^ Leakey, Richard. The Origin of Humankind, Science Masters Series. New York, NY: Basic Books. 1994: 87–89. ISBN 978-0-465-05313-1.
80.^ Jorde LB, Bamshad M, Rogers AR. Using mitochondrial and nuclear DNA markers to reconstruct human evolution. Bioessays. February 1998, 20 (2): 126–36. doi:10.1002/(SICI)1521-1878(199802)20:2<126::AID-BIES5>3.0.CO;2-R. PMID 9631658.
81.^ 引用錯誤:無效<ref>標籤;未為name屬性為1002/ajpa 351的引用提供文字
82.^ Wall, J. D.; Lohmueller, K. E.; Plagnol, V.. Detecting Ancient Admixture and Estimating Demographic Parameters in Multiple Human Populations. Molecular Biology and Evolution. 2009, 26 (8): 1823–7. doi:10.1093/molbev/msp096. PMID 19420049.
83.^ Green RE, Krause J, et al. A draft sequence of the Neandertal genome. Science. 2010 May 7;328(5979):710-22. PMID: 20448178
84.^ ^ Reich D, Green RE, Kircher M, et al. (December 2010). "Genetic history of an archaic hominin group from Denisova Cave in Siberia". Nature 468 (7327): 1053–60. doi:10.1038/nature09710. PMID 21179161.
85.^ Reich D ., et al. Denisova admixture and the first modern human dispersals into southeast Asia and oceania. Am J Hum Genet. 2011 Oct 7;89(4):516-28, PMID: 21944045 .
地球上生命的起源和演化
作者:李家維(演講人) 國立自然科學博物館、清華大學生命科學系
張志玲(文字整理) 《科學發展》特約文字編輯

圖說:「海口蟲」立體復原圖。
未來,我們的子孫會往哪兒發展呢?對於宇宙萬物、地球歷史、腳底下不安世界多些了解以後,某些科學家預測,未來的人類不是在地球上滅絕,就是要離開地球才能存活。
想要了解地球生命的起源和演化,總不免抬頭看看,這個宇宙到底如何。在這個超過一百五十億年的宇宙裡,曾形成無數星球,也曾幻滅許多星球。像我們銀河系這樣的星系可能有上千億個之多。若將範圍縮小,只看地球所在的銀河,類似我們太陽的星球,約有兩千億個;每個太陽旁邊,又帶了些行星。可以想像,這個宇宙有多少像地球一樣的環境可以孕育生命呢?所以說,生命遍布在整個宇宙的想法是可以預期的。
生命起源說各有論述
地球,大約在四十六億年前形成。過去二、三十年來,對於地球上生命起源的說法,一直在變,剛開始只是小幅度的改變,現在卻有較大膽的狂野想法。
過去的主流想法以「化學演化」來說明地球生命的起源。早期認為,地球生命開始於小分子、大分子到聚合物;從組成細胞、複製遺傳基因至細胞分裂繁殖,生命就開始了。但此場景究竟發生在什麼時候?產生了什麼樣的細胞?其實仍有很大的爭論。
其中一種想法是,生命可能來自於冰團。當海洋表面結成冰時,在冰晶縫隙裡,有機小分子聚成了大分子,生命於是開始。另一種想法是,受到月亮牽引潮汐的影響,海水蒸發,濃縮有機分子,岩石表面催化小分子的聚合,生命就這樣開始。最晚出現,大約一九七八年提出的想法認為,生命應從熱鍋開始:海底裂縫冒出的滾燙海水,最高超過攝氏300度,海水中夾雜很多特異的化學分子,這些化學成分和岩石作用後,不僅產生了海水裡的重要成分,生命也有可能從那裡開始。
生命跡象出現在各個角落
化學演化說在過去幾年開始鬆動,大家不再那麼信心滿滿。七年前的夏天,美國太空總署對外宣稱,在一塊火星隕石上發現生命跡象。原來,一位美國女科學家於一九八四年在南極冰層上撿到一塊石頭,經分析確定,它是來自火星的第 12 塊被檢驗出來的隕石。見到石頭上微小的形體,太空總署有了些想法,這些想法,其實在過去 20 年裡已有許多生物學家提出。
究竟是什麼呢?在北投、在黃石公園、在一些可以把蛋煮熟的熱泉裡,竟然住著能行光合作用的細菌,它們不只耐熱還嗜熱。此外,若向地下鑽個三千公尺,那裡見不到陽光,得不到有機物滋養,卻也住了很多細菌,它們吃、睡、繁殖都在那兒,是一群會利用自然界化學分子與氧化還原反應取得能量的細菌。
另有些細菌,在琥珀裡睡了二千萬年後還能被叫醒。這樣的場景,在地球上很多地方都找得到。例如美國加州大學洛杉磯分校沈育培教授,她在中國東北的黑土底下找到許多古蓮子,用碳-14 定年後發現,這些蓮子是依照唐、宋、元、明、清的順序躺在地下,其中最老的一顆,經鑑定已有 1,288 年歷史。若拿銼刀銼皮,讓蓮子吸水,再放入水裡三天,它不但萌芽,還能長出新的蓮苗來。
所以說,不只是細菌的生命堅毅,其他生物的生命也很堅毅。如果在高壓、高熱、高酸等極惡劣環境下生命仍可以存活,那麼,有什麼理由認為,這些生命不可以在星際間旅行呢?有什麼理由認為,它們不可以從一個星球散布到另外一個星球呢?
生命始於三十九億年前
這些堅毅的生命是從什麼時候開始的?科學界的說法不停地在修正,最近一次的說法是三十九億年前,那時候,天外流星仍不停地、猛烈地轟炸地球,在那擾攘不安的時代,生命已經開始。這個依據來自一些有機碳化合物的分子。
在寒冷的格陵蘭島上,某些磷酸鈣小砂礫裡包含著一些黑色的有機物質。雖然有機物質可以從自然界產生,也可以由生物合成,但是,分析當中的碳-12、碳-13比例後可知,這些黑色物質是由生物合成的。因為,只有生命可以擾動碳-12、碳-13 的比例。若這個定義正確,那麼,在三十九億年前磷酸鈣形成的時候,那些小砂礫已經包住了細菌的分子。如果生命曾出現在三十九億年前,也有可能不只出現一次。地球上的生命曾遭遇多次浩劫,人類則是災難倖存者的後代。
共生──演化中的重要策略
想要為生命痕跡尋找證據嗎?疊層石(stromatolite)是個好幫手。因為,疊層石是一種活石頭,在它的表面,有一群活著的細菌在那兒行光合作用。這些細菌,每天曬太陽、分泌膠質、將水裡的礦物質膠結住,也主動地把這些礦物質沉積下來。而石頭本身,就這麼一丁點一丁點地,一層層往上長。可以說,那些製造疊層石的生物,改變了地球空氣的組成,也決定了生命演化的方向。
地球就像其他行星一樣,在剛形成的時候沒什麼氧氣,只因氧是行光合作用的副產品,當氧氣隨著行光合作用的生物滋長且快速地增加以後,所做的第一件事,就是把地球給銹了。鐵,是地球上的第四大元素,在還原狀態時可溶於水,氧化後即沉積為鐵礦;現今煉鋼廠裡的鐵礦,大都是二十五億年前沉積的氧化鐵。沒錯,就在二十五億年前,疊層石生物產生的氧,把地球上的鐵都銹光了,之後,氧氣在大氣中開始侵害生命。
我們的祖先細胞因此面臨抉擇:滅絕,或是搬到沒有氧氣的地方居住。不過,聰明的祖先細胞選擇了「與細菌共生方式」存活下來。在此之前,細胞內沒有細胞核,所有化學反應都混在細胞質裡進行。當祖先細胞吞進原本獨立生活、又能利用氧氣的細菌後,開始發展「將不同反應放在不同細胞器官進行」的能力,既解決氧氣毒害問題,又強化了代謝協調的效率。細菌進入祖先細胞後,就再也無法和我們分開了,這是共生現象(symbiosis),是生命演化過程中非常重要的策略。
圓滾滾的動物胚胎
大約六億年到八億年前,至少有兩次,或者四、五次,地球凍成了冰球,每次凍結,就凍個千把萬年。地表上所有的水幾乎都成了冰,對生命而言,真是嚴苛的考驗,許多生命因此滅絕。
這現象一直維持到五億八千萬年前,地球回暖,大陸貴州附近變成溫暖的淺海。歷經兩、三億年後,貴州又逐漸變成陸地。四千萬年前,印度板塊與歐亞大陸板塊碰撞、擠壓,拱起了青康藏高原,也隆起了雲貴高原,於是,累積在貴州的三、四千公尺的岩石,開始面對幾千萬年的風化侵蝕,到最後,終於土崩瓦解,露出藏在五億八千萬年前那些溫暖淺海裡的生物紀錄。
在貴州瓮安縣有個五億八千萬年前的地層,裡面發現許多比小米還小的顆粒狀構造,圓滾滾的外形,大大小小有好多種。把它們拿出來用放大鏡看,就能看到上頭的裂縫,一個裂成二個、四個、八個、十六個……,到底是什麼呢?動物的卵和胚胎!放在電子顯微鏡下觀察時,可清晰見到每一個細胞的構造;磨成薄片後,還可見到細胞與細胞間的界線,而且,每一個細胞核都在,連蛋黃構造都保存著。
到底是什麼動物能在遠古的歲月裡生下這麼多蛋呢?剛開始時,研究人員把石頭磨成薄片,或從裡面挑出胚胎,然後一個個去辨認;有時候敲開它們,看看內部構造。但對如此精美難得的化石,任何破壞都是一種罪惡,看來,要分析石頭裡的結構,非得研發更先進的技術不可。
非破壞性分析研究
正巧新竹清華大學隔壁是國家同步輻射研究中心,如果把粒子加很高能量後,可導出平行、高解析度的X光,這就可以為化石拍照,希望能顯現化石內部的細微立體構造。後來發現這個辦法真不錯,於是尋訪世界各國的同步輻射中心,看誰可以把這項技術做得更好。
如今這種非破壞性分析的X光顯微術研究,已在美國、韓國、瑞士、臺灣等的同步加速器中進行。這些研究隱含著一個迷人的期待:十年以後,美國太空船從火星上帶回石頭,應該只有少數人有機會參與研究,到時候,誰有本事以非破壞性分析技術來看火星岩石裡面最細微的構造,誰就有機會爭取研究的優先權。
四月天的海底下熱鬧極了
從瓮安化石層一塊拳頭大的石塊裡,就可以找到上萬個動物胚胎,地球上還有這樣的場景嗎?其實在我們身邊就有。每年四月,月圓過後的第二天晚上,墾丁海底下的珊瑚都會集體產卵,無數的美麗卵泡,瞬間密布海水;不只珊瑚,很多無脊椎動物都在產卵,蚌殼把大量的精卵送出來,海膽、海星都在做這件事,四月天的夜晚,海裡頭熱鬧極了!
同樣場景,也可能出現在五億八千萬年前的瓮安,四月天,月圓後第二個晚上,海洋中的無脊椎動物正大量地釋放精卵。但災難突然來到,發育中的大量胚胎,很快地被殺死,再沖到附近一個有大量磷酸與鈣的海域,這些物質,又很快地把胚胎化成不朽的石頭。
地質學家習慣把地球的歷史切成兩段,一段稱為前寒武紀,另一段的開端就是古生代的第一紀──寒武紀(Cambrian)。過去一百多年來,寒武紀的開端(五億四千萬年前)被認為是動物出現的時候,然而,出土大量動物胚胎化石的貴州瓮安地層,卻比寒武紀的開始早了四千萬年。顯然地,在寒武紀之前,地球上早就出現了很多複雜的動物。
海口蟲──演化中的重要生物
大陸昆明東南 50 公里遠的澄江縣也有個特別地層。五億兩千萬年前,那兒是一片溫暖淺海,但不知怎麼地,大約每隔一百年,附近就有個災難。只要災難一來,大量細緻的粉沙,會快速地把住在淺海裡的生物掩埋下來;在極短時間內,脫水、壓扁,許多生物還來不及被細菌腐化,就被壓成扁扁的化石,所有的構造細節都留在岩石上。那兒有很大一片區域布滿了這種化石,這些化石總稱為澄江生物群(Chengjiang Biota)。
在滇池旁有一個叫做海口的小鎮,研究人員在一處不到四平方公尺的範圍內,找到三百多條小蟲子,取名海口蟲(Haikouella)。牠的肌肉已經分節,有消化道、尾巴、肛門與四對生殖腺,更重要的是,牠有一個脊索動物的特徵──軟的彈性構造。
脊椎動物的特質是脊索,在當時,我們的祖先沒有選擇堅硬的盔甲,反而長出背部的脊索以便彈性地應付這個世界;又因為捨棄盔甲選擇智慧,所以長出個大腦來。脊索與大腦,成為無脊椎動物與有脊椎動物的最大區別,而海口蟲兼具這兩種構造,牠被認為是生物演化過程中一個非常重要的環節,也是無脊椎動物演化成脊椎動物的典型過渡代表。
連續火山爆發古生代結束
古生代(the Paleozoic era)有很多紀,每一紀的結束,代表一次災難;每一次災難,都是生物滅絕的時候。到了古生代末期,陸塊逐漸聚集,海岸線消失,陸地中央變成沙漠。所以,地球變得很乾,有些生物耐不住乾旱而逐漸滅絕。
但是,有一件事澈底改變了這一切。大多數說法認為,現今的西伯利亞地方,在二億五千萬年前發生了連續的火山爆發,這個爆發大到無以描述,在二百五十萬平方公里的面積上,累積了四公里厚的岩漿,如果將它攤平到全世界,大概可以攤到七公尺厚。岩漿噴了多久?沒人知道,但必定噴出大量火山灰,遮天蔽日;噴出很多氧化硫,折射陽光,使地球冷卻;噴出大量毒氣;噴上天的氧化硫,遇水變成硫酸,落到地下變成酸雨。可以想像,那是場大浩劫!
結果,陸地上幾乎所有生物都死了,陸地上的大型動物超過三分之二消失;海洋裡,活下來的生物不到 5%;災難後的地層裡,不再有卷柏、木賊的孢子,取而代之的是腐化這個世界的真菌孢子。你能想像那個悲慘歲月嗎?滿地毛茸茸的真菌,正快速地腐化死亡的生物遺體。然而,危機也是轉機,又有很多新生物開始出現,古生代結束,中生代(the Mesozoic era)開始!
蟲鳴鳥叫恐龍是主角
中生代有些什麼呢?爬行生物飛上天,變成翼手龍;入了海,變成魚龍。最後,大到無以倫比,長度可到六、七十公尺,走起路來地都震了的恐龍變成主角。而中國東北的遼西地區,從那兒出土的帶羽毛恐龍化石,震驚全世界,使得長達 150 年的「鳥的祖先可能是恐龍」的爭論,暫時停火。
回溯一億兩千四百萬年前,遼西地區是一片大森林,布滿沼澤,花已經演化出來,蟲鳴鳥叫,生機盎然。依照人們尋找化石的經驗,天上飛的鳥通常不容易保存為化石,但在那兒,卻有上百種、幾萬個鳥化石,有些精美的標本上,每根羽毛都在。當時在那兒必定經常發生火山爆發,每次爆發,就在地層裡留下一層層火山灰,有些厚,有些薄,頻率相當高。
稍微想像一下:火山爆發,火山灰,快速侵蝕那個生態世界,鳥、獸受到驚嚇,爭先恐後地逃出林子,拼命往水邊跑。有些鳥,飛到一半,被後頭的火山灰、毒氣趕上,栽下來,掉到爛泥裡,掉到火山灰裡。火山灰的粉末極細緻,滲入鳥翅膀的每個空間,鳥羽毛被完整地保存下來,細菌都還來不及分解,整隻鳥就變成了化石。帶羽毛恐龍化石,想必也是如此形成的。
自從一九九六年,第一隻帶羽毛恐龍化石被發現後,直到現在,這兒又陸續發現十幾種帶羽毛恐龍。但是,為何世界上其他地方未曾發現呢?除恐龍種類不同外,應是遼西地區特殊的地質條件和埋葬條件,使得羽毛能夠清晰地保存下來。
植物開花生命共同演化
地球上有這麼豐富的色彩,這麼多種類的生物,能夠造就這麼多樣性的世界,都要歸功於植物開花。植物開花的目的不在吸引同伴,花蜜和花香是為動物製造的,一旦動物被吸引過來,就可幫忙傳播花粉,傳播種子。
昆蟲,在植物身上留下了蛋,蛋孵出來變成毛蟲,毛蟲開始啃食葉子;植物不喜歡,為了活下去,就做出毒汁、長出刺;蟲子要活下去,又演化新的能力,目的是要分解毒汁,閃躲刺。地球上生命的合作競爭,就這麼反覆發生,這就是生物學上所說的共同演化(coevolution)。
在很短時間內,共同演化造就了三十萬種以上的開花植物,和一百萬種,甚至一千萬種的昆蟲。在什麼時候,什麼場景下,哪一種植物在地球上開出第一朵花,是演化生物學家很想知道的答案。只是,花如此脆弱,在植物生命裡出現的時間這麼短暫,又怎能變成化石呢?因此,找到最古老的開花化石,是科學家極度渴望的事。
一九九八年,中國科學院南京地質古生物研究所孫革等人發現了遼寧古果化石,相關文章在英國《自然》雜誌發表,可惜化石上只有果子,沒看到花;之後中國地質科學院地質研究所的季強和孫華,在遼西發現保存完整的中華古果化石,相關報導在二○○二年《自然》雜誌上刊出。如今國際科學界已承認,古果化石是目前世界上已知最早的被子植物。
終於看到祖先的模樣
人類屬於「有胎盤哺乳動物」,對於有胎盤哺乳動物的研究,最近幾年有驚人的發現。在遼寧淩源市義縣一億兩千四百萬年前的地層內,季強和美國卡內基自然史博物館的羅哲西發現真獸類(有胎盤類)哺乳動物化石,並取名攀援始祖獸(Eomaia scansoria)。相關研究報告在二○○二年四月出刊的英國《自然》雜誌發表。
在過去,生物學家沒看過有胎盤哺乳動物祖先的模樣,古生物學家也只能從岩層裡篩出細碎的牙齒做研究。這次發現的攀援始祖獸化石,毛在、牙齒在、每根骨頭都在;牠的身長約八公分,體重約 25 公克,具有可以靈活攀援的肢骨特徵,屬哺乳綱(Mammalia)真獸類(Eutheria),是世界上已知的,有胎盤哺乳動物中最古老、最原始的物種。所有包括人類在內的現有胎盤哺乳動物,都是真獸類哺乳動物的後裔。
天崩地裂新生代來臨
對於中生代的結束,最盛行的說法是:六千五百萬年前,一塊十公里大的石頭,挾持著重量與速度,掉落在墨西哥海邊,瞬間,海水蒸發!這個石塊繼續往下鑽,碰撞到的岩石是碳酸鈣和硫酸鈣,碳酸鈣產生二氧化碳,汽化了;硫酸鈣產生二氧化硫,出去了。二氧化硫衝上了天,和水結合,變成酸雨落到地面;未結合的,就把陽光反射回去。石塊再繼續往下鑽,更底下是矽酸岩組成的花崗石,花崗石熔化成岩漿,噴到天空,冷卻後,變成黑色的黑曜石,在附近灑下一片玻璃雨……。
地球正在天崩地裂之際,海水則猛烈沖擊著海岸,變成海嘯,更在岸邊掀起四千公尺高的滔天巨浪。今天的德州,仍可看到一道三百公里長的石牆,可能是當時隨海嘯沖過來的石頭堆疊起來的。這樣的撞擊,或許又讓位在地球另一邊的印度發生地震,引發火山爆發。我們的地球,陷入了什麼樣的困境呢?有火山灰、毒氣體、酸雨、沒有光線、植物死亡、動物餓死、超過 80% 的海洋與陸地生物滅絕、中生代結束!少量倖存的生物,重新開始迎接新生代(the Cenozoic era)的來臨!
http://web1.nsc.gov.tw/ct.aspx?xItem=8038&ctNode=40&mp=1
宇宙的起源
宇宙起源于137億年前的宇宙大爆炸,形成了衆多的星系和恒星;
此後,輕重不同的100余種元素不斷地通過氫和氦熱核反應而逐漸形成。
在46億年前一次很偶然的超新星爆炸沖擊波的影響下,使由星際冷的固體塵埃和氣體所組成的原始太陽星雲運動速度加大,密度增大,發生凝聚而形成太陽以及圍繞它旋轉的星雲物質。
太陽系的起源(4.6Ga)
太陽周圍的固體與氣體物質,經過約5000萬年快速旋轉和冷凝
以固體顆粒(小園球狀的星子)爲中心,通過反複隨機碰撞吸積、
增大質量
逐漸形成太陽系的9大行星。
地球的形成
根據球粒隕石 年代測定,地球的形成和加大吸積的時期,推測主要發生在45.2~ - 44.8億年前。
地球總質量的絕大部分 (6×1027 g) 都是該時期吸積的。
後來的40億年內,地球質量只增加1025 g,相當于地球總質量的1/600。
也就是說地球平均每年要接受2.5×1016 g (即250億噸)的隕石物質。
早期的地球—地核
當時地球內部熱量比現代大得多,估計其放射性幅射能約比現代大10倍;
? 地球吸積(隕石撞擊)和圈層分異作用也可産生熱量,使物質分異與遷移,比重較大的鐵鎳物質大量聚集到地心附近,到43億年前,就基本上形成地核(約相當于現代地核的3/4)。
早期的地球—地幔、地殼
地幔基本上一直保持固體狀態。
下地幔成分相對均一,而上地幔存在著橫向與縱向上化學成分的不均一性,說明上、下地幔並沒有經曆過全部的熔融
分異、以達到平衡的演化過程。
而在地球表部,火山密布,同時還有相當數量的隕石撞擊,在40億年前逐漸形成了原始的大陸,使晶體結構較松散的
鋁矽酸鹽在地表聚集,造成了大陸地殼。
大陸地殼的形成和增長
陸殼的主要增生時期爲30~16億年即新太古—古元古代時期,現在所知的大陸地殼基本上都是該時期形成的。
在18億年以前的時期,由于地球內部的熱活動一直比較強烈,物質在垂直方向(地球半徑方向)上的分異、對流、遷移等作用都相當劇烈。
? 在18億年以後,地球內部變冷,岩石可塑性變小,使上地幔的頂部層位和地殼合成一個統一的岩石圈,其剛性程度加大。
大氣圈的形成
與此同時,由于岩漿活動所排出的氣體(CO2與H2O爲主,含少量N2)可能形成初步的大氣圈,形成與金星、火星相似的大氣圈。
大氣壓力相當于現代的300倍,溫度可達230℃左右,以後逐漸降溫,氣體中冷凝下來的水(富含Cl、SO4離子),就可覆蓋在幾乎整個地球表面。
水的形成
1998年Zahnle認爲地球上水主要由富含冰(固體的H2O)的慧星撞擊地球所致。
關于水的起源,留下的證據不足,也許兩種成因──岩漿排氣與慧星撞擊可能都同時存在。
一般認爲85%的大氣圈與水圈(海洋)是在44億年前形成的。
http://jwc.cugb.edu.cn/dizhi/cugb_jpkc/diqiu/jiaoan/p11.htm
板塊運動
板塊構造學說(Plate tectonics)主要在說明目前發生在地球上層的構造及解釋地震發生之原因。
地球的最外部為冷而硬的可移動之岩石,稱為岩石圈(lithosphere ),其厚度平均約100公里,岩石圈之下為軟流圈(asthenosphere)為黏度高的液體物質所組成,在高溫、高壓作用下而成可塑性,使岩石圈漂浮其上。
板塊構造的基本觀念是將岩石圈分成數個接近剛性之板塊,包括較大的歐亞板塊、美洲板塊、非洲板塊、印度洋板塊、太平洋板塊及南極洲板塊和數個較小之板塊(見附圖),板塊受到張力、壓力、重力及地函對流的作用,不同的板塊之間每年以數公分的相對速度緩慢移動,大部分的地震、火山及造山運動便由於相鄰板塊之互相作用而發生。
板塊交界處主要有三種型態:
(1)分離板塊交界處(divergent boundaries):代表地殼引伸拉裂的現象,在中洋脊(mid-ocean ridge)處相鄰的兩板塊互相分離,而產生新的岩石圈,其材料來自地函的上部,係經熔融作用而產生。地殼在這裡由於張力作用向兩側擴張延伸,沿著發散交界處常有地震發生,其震源深度多在100公里以內。
(2)聚合板塊交界處(convergent boundaries):在這交界處兩板塊相互碰撞,較重者插入較輕者之下方(約以30°~45°之傾角),使老的岩石圈消失而回到地函中,這插入的部分叫隱沒帶(subduction zone)。由於兩板塊間的相互磨擦,所以沿著隱沒帶可以不斷地發生地震而造成一地震帶,其震源深度可從很淺到大約700公里左右。台灣花蓮附近為歐亞大陸板塊和菲律賓海板塊之聚合板塊交界處所以地震非常頻繁。
(3)守恆板塊交界處(conservative boundaries):不產生新的岩石圈也不使岩石圈消失,相鄰兩板塊彼此橫向移動磨擦,而產生震源深度較淺之地震。台東縱谷斷層即為歐亞大陸板塊和菲律賓海板塊之守恆板塊交界處。
http://scman.cwb.gov.tw/eqv5/eq100/100/051.HTM地球的演化




| 代 |
纪 |
世 |
Ma |
| 显
生
宙 |
新
生
代 |
第 四 纪 |
全新世 |
1.65
23.5
65
135
205
245
295
360
410
435
500
540
1000
1600
2500
3800
|
| 更新世 |
晚 |
| 中 |
| 早 |
| 第
三
纪 |
新第三纪 |
上新世 |
| 中新世 |
| 老第三纪 |
渐新世 |
| 始新世 |
| 古新世 |
| 中
生
代 |
白 垩 纪 |
晚白垩世 |
| 早白垩世 |
| 侏 罗 纪 |
晚侏罗世 |
| 中侏罗世 |
| 早侏罗世 |
| 三 叠 纪 |
晚三叠世 |
| 中三叠世 |
| 早三叠世 |
| 古
生
代 |
二 叠 纪 |
晚二叠世 |
| 早二叠世 |
| 石 炭 纪 |
晚石炭世 |
| 早石炭世 |
| 泥 盆 纪 |
晚泥盆世 |
| 中泥盆世 |
| 早泥盆世 |
| 志 留 纪 |
晚志留世 |
| 中志留世 |
| 早志留世 |
| 奥 陶 纪 |
晚奥陶世 |
| 中奥陶世 |
| 早奥陶世 |
| 寒 武 纪 |
晚寒武世 |
| 中寒武世 |
| 早寒武世 |
| 元
古
宙 |
新元古代 |
NeoproterozoicⅢ |
|
Cryogenian |
|
Tonian |
| 中元古代 |
Stanian |
|
Ectasian |
|
Calymmian |
| 古元古代 |
Staitherian |
|
Orosirian |
|
Rhyacian |
|
Siderian |
| 太古宙 |
|
Archean |
| 冥古宙 |
|
|
| 资料来源:Gilles S.Odin and Chantal
Odin(1990) |
冥古代的地球
冥古代(Hadean)是指自地球形成至距今38億年前這段時期,有些科學家稱為地球的天文時期、或地球的前地質時期、或前太古代、或原太古代。
這一時期地球歷史包括原始地殼、原始陸殼的性質和形成以及原始生命的形式和出現等複雜的問題。
地球起源於46億年以前的原始太陽星雲。
經過微星的集聚、碰撞和擠壓使其內部變熱,以後則是放射性物質的衰變使地球內部進一步升溫,約在距今45-40億年前,當溫度上升到鐵的熔點時,大量融化的鐵向地心沉降,並以熱的方式釋放重力能,其能量相當於一千多次百萬噸級的核爆炸。大量的熱使地球內部廣泛融化和發生改變,逐步形成了分層結構,其中心是緻密的鐵核,熔點低的較輕物質則浮在表面,經冷卻形成地殼。
當時地表的溫度、大氣和水體的組分和性質可能還不具備生命產生的條件,因而也不會出現風化侵蝕等地質作用及其產物。那時地球岩漿活動劇烈,火山爆發頻繁,表面覆蓋著熔化的岩漿海洋。以後,隨著地球溫度的緩慢下降和冷卻,同時由於上述的分異作用,一開始就可能使氣體逸出,蒸發的氣體不斷上升,在空中又凝聚成雨落回地面,隨著不間斷的雨水的侵入,原始大氣圈和海洋誕生了。這時大氣圈中含有大量的二氧化碳,地球也被厚厚的雲層封鎖著,太陽光幾乎穿不透地球橘紅色的天空,海洋的溫度高於150 攝氏度。在這沸騰的海洋裏,孕育生命的各種元素在不斷積累。
元古代的地球
元古代(Proterozoic Era)開始於距今大約25億年以前,結束於距今大約6億年前的"生命大爆炸"。這一時期,現在的陸地在那時大部仍然被海洋所佔據,地殼運動劇烈,到了晚期,北方勞亞古陸和南方岡瓦納大陸的面積擴大了許多,出現了若干大片陸地。在我國,許多地區已經露出海面而成為陸地,而西藏的大部分仍然被海水佔據。
元古代晚期在我國被稱為震旦紀(Sinian Period),時間為大約從距今8億年以前到元古代結束,震旦是古代印度對我國的稱呼。在震旦紀,出現了全球性的大冰期,稱為震旦紀大冰期,是地球發展史上的三大冰期之一。冰川最盛時覆蓋了亞洲、歐洲、美洲、大洋洲的許多地區,有的地方冰層厚達千米。
元古代時期,海水裏的生命活動明顯地加強了,生物界由原核細胞形式演變為真核細胞形式,但演變的過程和時間還不清楚。這時細菌和藍藻開始繁盛,後來又出現了紅藻、綠藻等真核藻類。藻類在生長過程中粘附海水中的沉積物顆粒形成層紋狀結構物,稱作疊層石,疊層石是地球上最早的生物礁,出現於太古代而在元古代達到頂盛。除了藻類生物外,元古代結束前,海洋裏出現了一些如海綿等低等無脊椎動物。"元古代"的意思,就是原始時代。
太古代的地球
太古代(Archaeozoic Era) 是最古老的一個地質年代,開始於距今38億年前,結束於距今25億年前。這一時期,地球上是一片深淺多變的廣闊海洋,沒有寬廣的大陸,只有一些孤島--原始的陸地,稱為陸核。海洋裏分散著一些火山島,陸地上只有禿山,一片荒涼。太古代時,地殼(原始陸殼)的組成成分可能與上地幔更為接近,或者由於地殼厚度較不,幔源物質容易向上運動,出現大規模的火山噴發。因此,那時岩漿活動劇烈,火山噴發頻繁,經常出現煙霧漫天的景象。
太古代,我國的華北大部、東北部分地區和新疆西部的部分地區已經露出海面,成為陸地,其他地區還是一片汪洋。北方大陸區--勞亞古陸包括北美、東歐和西伯利亞,已形成許多原始的古陸。同樣,南方大陸--岡瓦納大陸包括非洲、印度、澳洲、南美和南極洲,也形成了許多的原始古陸。但是,這些大陸區的古陸都被廣寬的海水所分隔,彼此並不相連。
太古代早期,海水中逐漸形成了一種類似蛋白質的有機質,慢慢就成為最原始的生命體。大約在距今約34億年前,原始海洋裏出現了能夠進行光合作用的藍藻。雖然在早期就開始有藍藻等原核生物出現,但那時形成的岩石在漫長的時期內經過了深度的變質,因此保留下來的可靠的化石非常少。太古代是形成鐵礦的重要時代。
早古生代的地球
從古生代(距今6億年~2.5億年)開始,地球歷史的發展進入了一個新的發展時期。
古生代分為早古生代和晚古生代兩個代。
早古生代(距今約 6-4億年前),地球發生了規模巨大的加里東運動,地殼變動劇烈,許多地方反復上升和下沉,陸地面積逐漸擴大。這一時期,北方大陸各地區仍然彼此分離,南方各大陸則相對集中,但也不是完全相互連接,北美與南方大陸的相對位置愈晚愈趨於接近,其與歐洲之間也是互相接近。當時隔開北方各大陸與南方大陸的古地中海(特提斯海)比現代的地中海更為寬廣,而亞洲東部和南部與澳大利亞東部有密切的關係。早古生代結束時,我國的陸地與震旦紀的區域基本一致,但面積有所擴大,南方大部分地區出露海面成為陸地。
由於上述原因,早古生代的生物區,一是以北美和歐洲為中心,稱北大西洋區;二是自東北亞向南,包括東亞、東南亞直到澳洲,稱為亞澳太平洋區;三是逐漸分異發展的古地中海區,又以有廣海隔絕而分為南北兩個亞區。
早古生代又分為寒武紀、奧陶紀和志留紀三個紀。
寒武紀的地球
寒武紀(距今6--5億年前)是早古生代的第一個紀,地球開始爆發了全球性的加里東構造運動,地球各板塊互相碰撞、擠壓,火山噴發頻繁,南、北大陸發生多次反復升降,許多地區重新又被海水所淹沒。北方各大陸的相對位置較震旦紀時靠近,但互相並沒有連接;南方岡瓦納大陸逐漸連在一起,但並不是完全相連,且可能通過非洲與北方大陸相連通。
寒武紀,最主要的特徵是生物界的顯著繁盛和化石的大量保存。
早期,氣候乾旱,淺海分佈廣泛,門類眾多的海洋無脊椎動物突然之間出現在地球上,這是生物史上的第一次大發展,被稱為著名的“生命大爆發”事件。從此,地球上的生物開始了大發展時期,並且空前繁盛,海水裏出現了許許多多種動物,如三葉蟲、頭足類、珊瑚等。
奧陶紀的地球
奧陶紀(距今5--4.4億年前)是地球歷史上大陸地區遭受廣泛海侵的時代,是火山活動和地殼運動比較劇烈的時代,也是氣候分異、冰川發育的時代。從奧陶紀起,海生無脊椎動物真正達到繁盛的時期,也是這些生物發生明顯的生態分異的時期。在奧陶紀後期,各大陸上不少地區發生重要的構造變動、岩漿活動和熱變質作用,使得這些活動區的部分地區褶皺成為山系,從而在一定程度上改變了地殼構造和古地理輪廓。
科學家認為,奧陶紀時期,各大陸相對於兩極的位置和大陸之間的相對位置都曾發生過重要的改變。當時,西伯利亞中北部、加拿大北部的部分地區、中國北部和澳大利中西部都屬於乾熱氣候的地區;相反,北非的撒哈拉沙漠、南非開普地區曾經覆蓋著厚厚的冰層,屬於寒冷氣候地區。這說明,奧陶紀時,古南極在現在的撒哈拉沙漠以南,古北極位於南太平洋,古赤道恰好穿過西伯利亞中西部和中亞一帶,經加拿大西部向南太平洋岸南下。
志留紀的地球
到了志留紀(距今4.4--4億年前),由於加里東運動的影響,地殼運動的強度和古地理地貌都發生了比較顯著的變化。從志留紀中期以後,地球各大陸區普遍發生海退,陸地隆起、面積擴大,許多地區也發生褶皺成為山區,同時岩漿運動和熱變質作相對集中。
志留紀時期,北方各大陸逐漸靠近,但各大陸之間仍然被廣闊的海洋所分隔;而南方岡瓦納古大陸雖然各陸塊聯繫緊密,但它們之間並非一個整體,各陸塊之間還是有海水分隔,有些地塊之間還存在廣闊的海域。同時,分隔南北兩大古陸的古地中海逐漸縮小。
志留紀中期,生物界最著名的事件是生物從海洋向陸地進軍,開始征服大陸。由於志留紀中後期大面積海退,生物礁大規模出現,原來的海生魚類等逐漸適應半淡水生活,志留末期已有淡水生物、濱海半淡水動物和半陸物植物。同時,半陸生的裸蕨類進一步繁育,逐漸形成泥盆紀的真正陸生植物群。
晚古生代的地球
早古生代末,北美大陸與古歐洲大陸經過加里東運動而拼合在一起。因此,在晚古生代(距今4-2.5億年前)開始的時候,全球存在四個相互分離的巨型大陸板塊:古歐美大陸、古西伯利亞大陸、古中國大陸和岡瓦納大陸。在晚古生代時期,地殼又發生海西構造運動,使這些大陸之間發生了一系列的碰撞、拼合,終於在二疊紀末(距今2.5億年前)形成了統一的聯合泛大陸--盤古大陸。
在晚古生代,特別是石炭、二疊紀,全球的氣候分帶日益明顯,岡瓦納大陸在石炭晚期至二疊早期時處於南半球的高緯度區內;在低緯度的熱帶和亞熱帶淺海地區則常常形成大型的生物礁和介殼灘。陸地上溫暖濕潤的地區,陸生植物的異常繁茂,常常形成大量煤層,而乾旱地區則形成鹽類礦產。受氣候帶和地理隔絕的影響,晚石炭至二疊紀的植物群具有明顯的植物地理分區。
泥盆紀、石炭紀早期,全球的生物分區不明顯,說明氣候分異較弱。岡瓦納大陸上僅有局部地區有冰川活動,晚石炭世、早二疊世早期,岡瓦納大陸進入冰川活動的全盛時期,冰磧物幾乎遍佈岡瓦納,全球氣溫顯著下降,生物的緯度分異效應相應增強,出現類似現代地表生物緯度分異現象。早二疊世晚期起,氣候逐漸轉暖,溫暖氣候帶向南北擴展 。
泥盆紀的地球
泥盆紀(距今4-3.6億年前)是晚古生代的第一個紀,從距今4億年前開始,延續了4000萬年之久。由於早古生代加里東運動影響的結果,同時,從泥盆紀開始,地球又開始發生了海西運動。因此,泥盆紀時許多地區升起,露出海面成為陸地,古地理面貌與早古生代相比有很大的變化。
在北方勞亞大陸,許多原來被海洋佔據的地區升出海面,成為陸地,而原為陸地的地區則重新凹陷,又被海水佔據。泥盆紀起,北美板塊與俄羅斯板塊已經連成一片大陸,歐洲北部成為山系,並在波羅的板塊連成一片大陸;而南歐仍然為寬廣的海洋所佔據,東歐俄羅斯板塊大部分地區發生凹陷,重新被海水佔據;新疆北部、內蒙古及興安嶺地區在泥盆紀時期仍為分隔華北、塔里木和西伯利亞板塊的古亞洲多島洋。但是,勞亞古陸內各大陸之間並沒有相連,它們之間仍然被廣闊的海洋所分隔。
在南方的岡瓦納大陸,各大陸的位置互相間靠得更近,但還沒有形成統一的整體。
中國泥盆紀的古地理面貌受加里東影響,與早古生代相比,古地理面貌發生了重大變化,大部分地區上升為陸地,華南和華北板塊之間的秦嶺洋明顯收縮。因此,在泥盆紀時期,華北和華南板塊呈現相隔不遠而又獨立發展的局面。華北、塔里木和西伯利亞板塊之間仍被古亞洲洋所分隔。揚子板塊與岡瓦納板塊及羌塘地塊之間則被古特提斯洋分隔。昆侖山、滇西、西藏等地仍為古特提斯海洋所佔據。
泥盆紀時,生物界發生了從海洋征服大陸的巨變,從這一時期起,生物才開始由海洋向陸地發展。
加里東運動完結後,形成全球範圍的海域縮退,其結果是陸地面積進一步擴充了,地形起伏複雜,氣候變得乾燥炎熱。在泥盆紀早期,適宜這種環境的裸蕨植物和原始的石鬆類為主獲得了迅速發展,逐漸佔據了遼闊的大地,地球開始披上了綠裝;泥盆中期出現了楔葉和前裸子植物,泥盆紀晚期,石鬆和真蕨類形成的森林已經初具規模,這為陸生生物的發展準備了條件。事實上,部分植物擺脫海洋登陸,客觀上為生物界的發展奠定了條件,是生物演化史上的重大事件。泥盆紀最引人注目的是是魚類的進步,原始的甲胄魚類和真正的有頜魚類都非常繁盛,被稱為"魚類的時代"。盾皮魚中的恐魚可超過十米,是海洋中的統治者。晚泥盆晚期,水體逐漸乾涸,促使部分魚類--古總鰭魚類,不得不用鰭在泥沼中爬行,當肉鰭演變成四肢,使它們能夠爬到樹林中去時,一個新的門類出現了,這就是兩栖類。
最早的兩棲動物是魚石螈。
那時的魚類都生活在淡水或濱海三角洲半醎水中,早期的魚類比較原始,還沒有上、下頜的分化,後來才出現較進步的類型。另一方面,海洋中的無脊椎動物仍然統治著那裏的世界,腕足類、珊瑚、層孔蟲、苔蘚蟲、雙殼類、牙形刺等生物在大洋中競相發展,其中腕足類是非常引人注目的一類生物。腕足類屬於底棲固著型生物,軟體由兩瓣殼所保護,此外,還有一個用於支撐和固定身體的肉莖。它們在喜歡固定生活在安靜的海底,與世無爭。
石炭紀的地球
石炭紀(距今3.6--2.9億年前)是地質歷史上最早的世界性重要的成煤時期,也是地殼運動比較活躍和氣候分異明顯的時代。當時地球上首次出現大規模森林,不但廣布於濱海低地,同時也延伸至大陸內部。石炭紀中、晚期的地殼運動使許多地區出現大型拗陷盆地,為世界規模的聚煤期提供有利的古地理條件。北方勞亞大陸,早古生代時北美和南方岡瓦納大陸的位置是彼此分離的,但從石炭紀到三疊紀卻互相連接。根據陸生脊椎動物的分佈情況,石炭紀時北美東部與西歐之間也存在廣泛的陸地聯繫。早石炭世末期,西歐發生了強烈的造山運動,引起萊茵海海西地區皺起成山,而其邊緣則凹陷。東歐俄羅斯板塊石炭紀時海侵範圍比泥盆紀時更廣,當時的海岸線達到莫斯科附近。當時,西伯利亞板塊的通古斯和庫茲涅茨克屬於溫帶潮濕氣候,哈薩克斯坦西部已進入乾旱氣候帶,而再南的古地中海則已屬於赤道附近的炎熱潮濕氣候。
南方岡瓦納大陸,由於海西運動的加劇,普遍出現大規模的冰川,到晚期達到高峰,並且冰川活動一直延續到二疊紀。同時,岡瓦納大陸在石炭紀晚期,形成了一個統一的岡瓦納大陸,大陸冰蓋的中心大體接近古南極位置。石炭紀是地質歷史中冰期和間冰期交替出現的時代,因此海平面升降相對頻繁。伴隨著海陸變遷和陸地森林植被的大量出現,石炭紀成為中國地質歷史中第一次重要的成煤時期。中國石炭紀的古地理面貌是泥盆紀的繼續和發展:華北--柴達木板塊與華南板塊之間的秦嶺小洋盆繼續存在。華北、塔里木和西伯利亞板塊間的古亞洲洋內部發生了重要的造山運動,許多地區窿起成為高山。華南部分地區,在泥盆紀末期地殼上升,海水退去,但到了石炭紀初又開始新的海侵,中、晚期海侵範圍更進一步擴大,從而導致了華南板塊的東、南緣發生新的分裂,西緣的古特提斯海進一步擴大。
石炭紀時,氣候比現在要炎熱許多、且潮濕,這使得真蕨類迅猛地繁榮起來,在溫暖潮濕的地方,到處都長滿鬱鬱蔥蔥的沼澤森林,兩棲動物稱雄大地,林間飛著許多有翅昆蟲,昆蟲中有體型巨大古蜻蜓。石炭紀早期,地球上的植物群比較一致,都是以石鬆--楔葉為主的擬鱗木植物群;到了晚期,植物群分化為熱帶性質的歐美植物群和溫帶性質的安加拉植物群。由於森林過盛,這個時期留下了大量的植物遺骸--煤,因此石炭紀又稱造煤時期。
石炭紀中、晚期是脊椎動物演化史上又一個具有重要意義的飛躍階段,這就是原始爬行動物的出現。石炭紀森林中的兩棲動物主要是迷齒類的始螈,到石炭中、晚期,出現了最早的爬行動物。已知最早的爬行動物是--林蜥。
二疊紀的地球
二疊紀(距今2.9--2.5億年前)是古生代的最後一個紀,也是地殼運動較為活躍的時期,世界各許多不穩定的地區窿起成為高峻的山系,而穩定的地區則出現大型內陸盆地,總趨勢是陸地面積擴大和淺海面積顯著縮小。
到了晚期,北方勞亞大陸和南方的岡瓦納大陸相互聯合在一起,地球上出現了一個互相連結而又南北對峙的統一大陸--聯合古大陸或稱潘加亞泛大陸,標誌著海西構造運動結束。
二疊紀是生物界顯著變革時期,大陸面積的擴大和氣候環境的多變,使地球的氣候分帶現象明顯,出現了許多植物區。裸子植物數量增加,在植物界佔據了主導地位,除了種子蕨和科達樹外,還出現了松柏、蘇鐵和銀杏。在二疊紀時,迷齒類中的塊椎類發展成為佔優勢的兩棲動物,爬行動物開始輻射演化,除了原始的杯龍類外,似哺乳爬行動物達到全盛,早二疊紀的似哺乳爬行動物是盤龍類,其中的異齒龍是當時陸地上的頂級掠食者,晚二疊紀時非常接近哺乳動物的獸孔類佔領了大地,其中的巨獸是古生代陸地上出現過的最大的動物。
到大約二億年五千萬年前,發生了大規模的生物集群滅絕,很多生物退出歷史舞臺,這次滅絕事件與白堊紀末期恐龍的滅絕事件相比,有過之而無不及,是地球歷史上最大規模的生物滅絕事件,古生代從此結束。
中生代的地球
中生代是地球歷史上最引人注目的時代。
三疊紀中期,聯合的泛大陸達到了鼎盛時期,三疊紀末的印支運動導致古特提斯海的封閉和新特提斯海的擴張,聯合泛大陸開始出現開裂的跡象,環太平洋帶從晚三疊世開始活動,侏羅紀、白堊紀以聯合泛大陸的分裂解體、以及大西洋、印度洋等新海洋出現為特徵。
從晚侏羅紀開始,海洋板塊與周圍大陸板塊的碰撞俯衝、擠壓作用導致亞洲東部和美洲西部陸緣區發生重要的環太平洋構造運動,形成規模宏大的環太平洋造山帶。中生代時期最重要的地史特徵是泛大陸的解體,尤以白堊紀時岡瓦納大陸的分裂飄移最為顯著,從而導致的新洋盆形成和老洋盆的衰退,全球大陸的構造古地理格局發生深刻的改變。
中生代,脊椎動物開始全面繁榮並出現了一些最令人不可思議的物種--恐龍。爬行動物在海、陸、空都佔據統治地位,因此中生代又被稱為“爬行動物時代”。但在白堊紀末期,稱霸地球達1.6億年的恐龍卻大規模滅絕了。
同時,在侏羅紀末期出現了真正的鳥類和哺乳動物;植物界中,開花結果的被子植物也同時出現,並逐漸取代裸子植物。從此地球一改過去的面貌,到處都是鳥語花香。
中生代可劃分為三疊紀、侏羅紀和白堊紀。
http://www.teacher.aedocenter.com/mywebB/NewBook-4/HG-16.htm
文章定位: