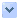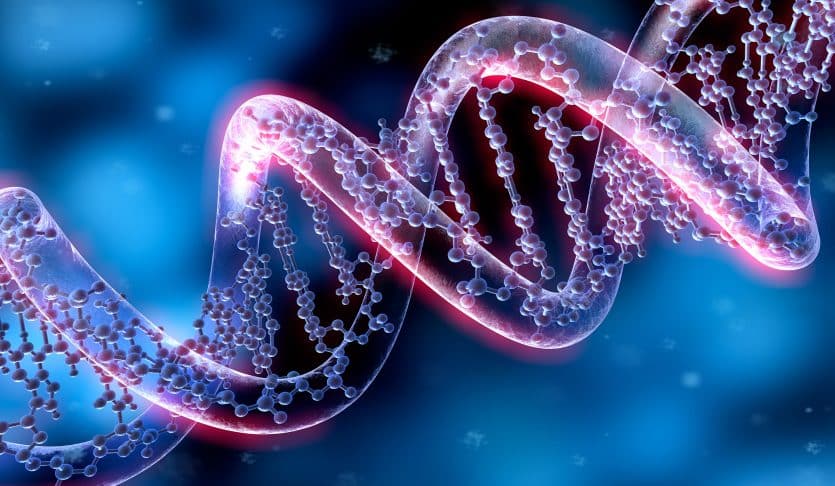
雜交種雜種的配子包含一個親本種(C)的基因組,而不是親本(紅色和綠色)染色體的所有可能組合(B)。A-體細胞。
*** 遺 傳 學 ***
遺傳學是研究生物體的遺傳和變異的科學,是生物學的一個重要分支。史前時期,人們就已經利用生物體的遺傳特性通過選擇育種來提高穀物和牲畜的產量。而現代遺傳學,其目的是尋求了解遺傳的整個過程的機制,則是開始於19世紀中期孟德爾的研究工作。雖然孟德爾並不知道遺傳的物理基礎,但他觀察到了生物體的遺傳特性,某些遺傳單位遵守簡單的統計學規律,這些遺傳單位現在被稱為基因。
基因位於DNA上,而DNA是由四類不同的核苷酸組成的鏈狀分子,DNA上的核苷酸序列就是生物體的遺傳資訊。天然DNA以雙鏈形式存在,兩條鏈上的核苷酸互補,而每一條鏈都能夠作為模板來合成新的互補鏈。這就是生成可以被遺傳的基因的複製方式。
基因上的核苷酸序列可以被細胞轉譯以合成蛋白質,蛋白質上的胺酸序列就對應著基因上的核苷酸序列。這種對應性被稱為遺傳密碼。蛋白質的胺酸序列決定了它如何摺疊成為一個三維結構,而蛋白質結構則與它所發揮的功能密不可分。蛋白質執行細胞中幾乎所有的生物學進程來維持細胞的生存。DNA上的一個基因的改變可以改變其編碼的蛋白質的胺酸,並可能改變此蛋白質的結構和功能,進而對細胞甚至整個生物體造成巨大的影響。 雖然遺傳學在決定生物體外形和行為的過程中扮演著重要的角色,但此過程是遺傳學和生物體所經歷的環境共同作用的結果。例如,雖然基因能夠在一定程度上決定一個人的體重,人在孩童時期的所經歷的營養和健康狀況也對他的體重有重大影響。

肌紅蛋白3D結構

DNA,遺傳的分子基礎。每一條DNA鏈都是由核苷酸連接而形成,兩條DNA鏈上的核苷酸鹼基互相配對形成螺旋梯結構,即DNA雙螺旋。

遺傳學的奠基者格里哥·孟德爾
遺傳學史
雖然遺傳科學開始於格里哥·孟德爾在19世紀中期的工作(包括實驗和理論),但其他一些關於遺傳的理論研究則早於孟德爾。在孟德爾時期,一種比較流行的理論——「混合遺傳」(blending inheritance)提出:個體的遺傳特徵是來自於其父母的特徵的混合平均值。孟德爾的工作則否定了這一理論,他的結果顯示遺傳特徵是由不同基因綜合表現的結果而不是連續的混合。當時的另一種得到人們支持的理論——「用進廢退說」(又稱為「獲得性遺傳」)提出生物經常使用的器官逐漸發達,不使用的器官逐漸退化,並且這種後天獲得的性狀是可以遺傳的。這一由拉馬克所提出的理論後來被證明是錯誤的,因為通常個體的經歷並不影響它們的基因,也就不會遺傳給下一代。
(註:但近年來對於表觀遺傳現象的研究發現,由於不同的經歷而引起的不同的性狀在一些情況下是可以被遺傳的,雖然這些發現並不能證明拉馬克的理論。)
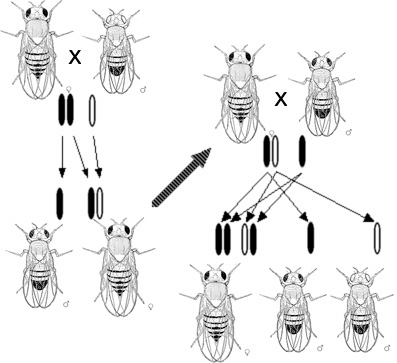
莫耳根對果蠅的白眼突變的性相關遺傳的觀察結果示意圖。莫耳根根據這一結果提出了基因是位於染色體上的假說。
古典遺傳學
現代遺傳學的奠基者是格里哥·孟德爾,一個奧地利修道士和科學家。他致力於研究植物的遺傳現象。1865年,他的論文《Versuche über Pflanzenhybriden》(《植物雜交實驗》)發表在布爾諾的自然研究學會(Naturforschender Verein)上。在論文中,他展示了豌豆在雜交實驗中所表現的遺傳規律並以數學關係加以描述。雖然這種遺傳規律只能在表現類型數量很少的情況下才能被觀察到,但是孟德爾的工作顯示遺傳是顆粒性的(顆粒遺傳,與混合遺傳相對,所謂的顆粒相當於現在為人們所知的基因),而不是混合性或獲得性的,並且許多性狀的遺傳規律可以通過簡單的規則和比率來解釋。
孟德爾工作的重要性並沒有得到廣泛的理解,直到他逝世之後的1890年代,當雨果·德弗里斯遇到相同的情況時才重新發掘出他的研究結果。作為孟德爾理論的支持者,威廉·貝特森(William Bateson)提出了genetics(「遺傳學」)這一名詞。1906年,倫敦召開的第三次國際植物雜交大會上,在貝特森的提議下,genetics這一單詞得以廣泛使用於描述關於遺傳的研究。
在重新評價孟德爾的工作後,科學家們試圖確定細胞中的哪一種分子是遺傳物質。1910年,基於對果蠅的性連鎖白眼突變的觀察結果,托馬斯·亨特·莫耳根提出基因位於染色體上。 1913年,他的學生Alfred Sturtevant利用遺傳連鎖的現象顯示了基因是在染色體上呈線性排列的。
分子遺傳學
雖然莫耳根等人的工作使得人們認識到基因是位於染色體上,但染色體是由蛋白質和DNA共同組成的,研究者們依然不知道哪一種物質才是遺傳物質。1928年,弗雷德里克·格里菲斯發現了轉化現象(參見格里菲斯實驗):死亡的細菌可以將遺傳物質「轉化」到其他依然活著的細菌內。16年後的1944年,奧斯瓦爾德·埃弗里、科林·馬克聊德(Colin McLeod)和馬克林·馬克卡提(Maclyn McCarty)鑑定出進行轉化的物質是DNA。
1952年,赫希-蔡斯實驗再次顯示是DNA(而不是蛋白質)才是感染細菌的病毒的遺傳物質,從而進一步證明了DNA是遺傳資訊的攜帶者。
1953年,利用羅莎琳·富蘭克林對DNA進行的X射線晶體學的研究成果,詹姆斯·杜威·沃森和弗朗西斯·克里克成功解析了DNA的雙螺旋結構。他們所提出的雙螺旋模型包含有兩條DNA鏈,鏈之間通過核苷酸上的鹼基配對,從而形成一個類似於旋轉梯子狀的結構。DNA結構顯示了遺傳資訊存在於每條DNA鏈的核苷酸序列中。這一結構也提示了一種簡單的DNA複製方法:兩條配對的DNA鏈分開後,新的配對鏈可以根據舊鏈上的序列來搭建。
雖然DNA結構顯示了遺傳的進行方式,但人們依然不知道DNA是如何影響細胞行為的。隨後的多年時間中,科學家們試圖了解DNA是如何控制蛋白質的製造過程。科學家發現細胞利用DNA作為模板來生成配對的信使RNA(RNA是一種類似於DNA的分子)。信使RNA上的核苷酸資訊就被用於生產蛋白質上的胺酸序列;這種由核苷酸序列到胺酸序列的轉譯是根據遺傳密碼的規則進行的。
隨著遺傳的分子機制的揭示,大量的研究成果不斷湧現。其中,一個重要的發展是弗雷德里克·桑格於1977年提出的鏈終止DNA測序法,這一方法使得科學家們可以閱讀DNA分子上的核苷酸序列。
1983年,凱利·穆利斯發展出了聚合酶鏈鎖反應技術,從而為從混合物中分離和擴增特定的DNA提供了一個快捷而靈敏的方法。經過人類基因組計劃和同時的競爭者塞雷拉基因組(一個私人贊助的基因組計劃)的努力,人類基因組的測序在2003年得以基本完成。

一個旁氏表(Punnett square),顯示了帶有兩種花色基因(紫花B和白花b)的豌豆雜交後的結果。
遺傳的特點
不連續遺傳和孟德爾定律
在最基本的水平上,生物體中的遺傳表現為離散性狀,即基因型。這種特點是由孟德爾首次觀察到,他研究了豌豆中遺傳性狀的分離現象。在研究花色的實驗中,孟德爾觀察到豌豆花的顏色只有兩種:紫色和白色,卻沒有任何一朵顯示出兩種顏色的中間色。這些來自於同一基因卻不同且離散的版本被稱為等位基因。
在豌豆的例子中,每一顆豌豆都含有一個基因中的兩個等位基因,並且子代可以從父母分別繼承其中的一個等位基因。許多生物,包括人類,都有這樣的遺傳規律。具有相同的兩個等位基因的生物體被稱為純合體,而具有不同等位基因的生物體則被稱為雜合體。
一個給定的生物體的等位基因的組合形式就是該生物體的基因型,而對於這種組合所表現出來的性狀就是該生物體的表現型。當生物體是雜合體時,常常有一個等位基因是顯性基因,顯性基因決定了生物體的表現型,而另一個基因就被稱為隱性基因,其性狀在顯性基因存在時不會被表現出來。有一些等位基因沒有完全的顯性,即「非完全顯性」,其表現為一種中間狀態的表現型,或者兩個等位基因無顯隱性之分可以同時表現出對應性狀。
當一對生物體繁殖後代時,它們的下一代隨機地繼承父母的兩個等位基因中的一個。這些對於離散遺傳和等位基因分離的觀察結果被總結為孟德爾第一定律(分離定律)。

遺傳系譜圖,可以幫助追蹤性狀的遺傳規律。
注釋和圖解
遺傳學家利用注釋和圖解來描述遺傳。一個基因可以用一個或幾個字母來表示,並且用大寫字母表示顯性基因,小寫字母表示隱性基因。「+」常常被用於標識一個基因的正常非突變的等位基因。
在雜交實驗中(特別是在討論孟德爾定律時),父母代被標示為「P」代,其下一代標示為「F1」(第一代)。F1代的子代就被稱為「F2」(第二代)。可以用於預測雜交結果的常用圖解是龐氏表(Punnett square,又稱為「棋盤法」)。
在研究人類遺傳疾病時,遺傳學家常常利用系譜圖來展示遺傳性狀。這些圖表將一個性狀的遺傳關係以家族譜(family tree)的形式表現出來。

人類的高度是一個複雜的遺傳性狀。來自法蘭西斯·高爾頓的1889年的資料顯示後代高度之間的關係是一個父母平均高度的方程。而從中計算的後代高度與真實值之間依然存在偏差,表明環境對這一性狀也有重要影響。
多基因的相互作用
生物體具有成千上萬個基因,並且在有性繁殖的生物中,這些基因的分類是互相獨立的。這就意味著對應豌豆的黃色或綠色的色彩等位基因的遺傳與紫色或白色的花色等位基因的遺傳是不相關的。這種現象被稱為孟德爾第二定律(又稱為「獨立分配定律」),即來自父母的不同基因的等位基因被隨機抽取來組成具有多種組合結果的子代。有一些基因不是獨立歸類的,這也就解釋了遺傳中的遺傳連鎖現象(具體內容見後)。
不同的基因常常能夠通過某種方式來影響同一種性狀。例如,在藍眼瑪莉(Blue-eyed Mary,一種植物)中存在一種能夠決定花色為藍色或洋紅色的基因以及一種能夠決定花是否有顏色(即白色或有色)的基因;當一株該種植物含有兩個白色等位基因(決定花色的第二種基因的兩個等位基因),則無論第一種基因所帶的顏色基因的等位基因為何,它的花色都為白色。這種基因之間的作用關係被稱為上位性或異位顯性(epistasis),即第二種基因位於第一種基因的上位。
許多性狀沒有明顯的可區分的特點(例如不同的花色),而表現為連續性的特點(如人類的身高和膚色)。這些複雜的性狀是來自於許多基因共同作用的結果。這些基因的影響作用在不同程度上是由一個生物體所經歷的環境所介導的。生物體的一個基因對於一種複雜性狀產生的影響的程度被稱為遺傳力。對一種複雜性狀的遺傳力的測量是相對的:環境的變化性越大,環境對於性狀變化的影響力也就增強,而基因對於性狀變化的影響力也就越小(表現為遺傳力降低)。例如,作為一種複雜性狀,美國人身高的遺傳力為89%;而在奈及利亞,由於人們所獲得的食物和醫療保健的差異性較大(即較大的環境變化性),其身高的遺傳力僅為62%。

DNA分子結構,雙鏈之間通過氫鍵排列形成鹼基配對。

細胞中染色體的結構。
遺傳的分子基礎 DNA和染色體
基因的分子基礎是去氧核糖核酸(DNA)。DNA由核苷酸相互連接而形成的鏈分子,其中的核苷酸有四類:腺苷酸(A)、胞嘧啶(C)、鳥苷酸(G)和胸腺嘧啶(T)。遺傳資訊就儲存在這些核苷酸序列中,而基因則以連續的核苷酸序列存在於DNA鏈中。病毒是唯一的例外,有一些病毒利用核糖核酸(RNA)分子來代替DNA作為它們的遺傳物質。
DNA通常以雙鏈分子的形式存在,並捲曲形成雙螺旋結構。DNA中的每一個核苷酸都有自己的配對核苷酸在相反鏈(對應另一條鏈)上,其配對規則為:A與T配對,C與G配對。因此,雙鏈中的每一條鏈都包含了所有必要的遺傳資訊。這種DNA結構就是遺傳的物理基礎:DNA複製通過將互補配對的雙鏈分開並利用每條鏈作為模板來合成新的互補鏈,從而達到複製遺傳資訊的目的。
不同基因沿著DNA鏈線性排列形成了染色體。在細菌中,每一個細胞都有一個單一的環狀染色體;而真核生物(包括動物和植物)則具有多個線形染色體。這些染色體中的DNA鏈常常會非常長;例如,人類最長的染色體的長度大約為247百萬個鹼基對。染色體DNA上結合有能夠組織和壓縮DNA並控制DNA可接觸性的結構蛋白,從而形成染色質;在真核生物中,染色質通常是以核小體為單位組成,每一個核小體由DNA環繞一個組織蛋白核心而形成。
一個生物體中的全套遺傳物質(通常包括所有染色體中DNA的序列)被稱為基因組。 僅含有一套染色體的生物被稱為單倍體生物;大多數的動物和許多植物為雙倍體生物,它們含有兩套染色體(姐妹染色體),即含有每個基因的兩個拷貝。一個基因的兩個等位基因分別位於姐妹染色體上的等同的基因座,每一個等位基因遺傳自不同親本。
性染色體是雙倍體生物中染色體的一個例外,它是許多動物中的一種特異化的染色體,決定了一個生物體的性別。在人類和其他一些哺乳動物中,性染色體分為X和Y兩類。Y染色體只含有很少量的基因,能夠觸發雄性特徵的發育;而X染色體與其他染色體類似,也含有大量與性別決定無關的基因。雌性具有兩個X染色體,而雄性具有一個Y染色體和一個X染色體。這種X染色體拷貝數的差別是性連鎖的遺傳病具有特殊遺傳規律的原因。

華爾瑟·弗萊明1882年的著作《細胞受質、細胞核以及細胞分裂》中描述真核細胞分裂的插圖。染色體被複製、聚集和組織。隨後,當細胞分裂開始時,複製後的染色體分別被分離進入兩個子細胞中。
無性繁殖和有性繁殖
當細胞分裂時,它們的基因組被複製產生兩份拷貝,每個子細胞繼承其中的一份。這一過程被稱為有絲分裂,它是繁殖的最簡單形式,也是無性繁殖的基礎。無性繁殖也能夠發生在多細胞生物體中,子代從單一親本處繼承其基因組,即子代與親本具有等同的基因組。這種子代與親本在遺傳上等同的現象被稱為複製。
真核生物常常利用有性繁殖來產生後代,其後代含有分別遺傳自不同親本的混合的遺傳物質。有性繁殖的過程是一個介於基因組單拷貝(單倍體)和雙拷貝(雙倍體)之間的一個轉換過程。雙倍體生物通過不複製DNA的分裂來形成單倍體,所生成的單倍體子細胞含有每對姐妹染色體中的任意一個。兩個單倍體細胞融合併將各自的遺傳物質組合在一起來重新生成一個含配對染色體的雙倍體細胞。多數動物和許多植物在它們的生命周期的多數時間內是雙倍體,只有生殖細胞為單倍體形式。
雖然細菌沒有單倍體/雙倍體的有性繁殖方式,它們也有許多獲得新的遺傳資訊的手段。一些細菌能夠發生接合,將一小段環狀DNA傳遞到另一個細菌細胞內。細菌還能夠從環境中攝入DNA片斷,並將之整合到自己的基因組中,這種現象被稱為轉化。這些進程導致了基因的水平轉移,即無親緣關係的生物體之間進行遺傳資訊的傳輸。
重組與連接
染色體的雙倍體使得位於不同染色體的基因在有性繁殖期間能夠獨立地分配(參見獨立分配定律),並通過重組形成新的基因組合。在同一條染色體上的基因理論上不會發生重組,但通過染色體互換則可以達到。在互換過程中,染色體交換DNA片段,有效地將染色體之間的等位基因重新分配。染色體互換通常發生在減數分裂期間(旨在生成單倍體細胞的一系列的細胞分裂過程)。
染色體上兩個給定位點之間發生染色體互換的可能性與這兩個位點之間的距離相關。對一個任意長的距離,由於互換的可能性足夠高,使得相隔該距離的兩個基因的遺傳無相關性。對於兩個接近的基因,由於互換的可能性較小,則基因之間會發生遺傳連鎖,即這兩個基因的等位基因趨向於被一起遺傳。一系列基因之間的連鎖數量可以被組合在一起構成一個線性的連鎖圖譜來描述染色體上基因的排列順序。

染色體之間的互換和雙互換。

遺傳密碼:DNA通過信使RNA作為中間載體編碼蛋白質。
基因表現
遺傳密碼
基因通常是通過生成所編碼的蛋白質(執行細胞中大多數功能的複雜的生物大分子)來表現它們的功能性影響。蛋白質是由胺酸所組成的線性鏈,而基因的DNA序列(通過RNA作為資訊的中間載體)被用於產生特定的蛋白質的胺酸序列。這一過程的第一步是由基因的DNA序列來生成一個序列互補的RNA分子,即基因的轉錄。
通過轉錄產生的RNA分子(信使RNA)被用於生產相應的胺酸序列,這一轉換過程被稱為轉譯。核酸序列中的每一組三個核苷酸組成一個密碼子,可以被轉譯為20種出現於蛋白質中的胺酸中的一個,這種對應性被稱為遺傳密碼。這種資訊的傳遞是單一方向性的,即資訊只能從核苷酸序列傳遞到胺酸序列,而不能從胺酸序列傳遞迴核苷酸序列,這一現象被弗朗西斯·克里克稱為分子生物學中心法則。
特定的胺酸序列決定了對應蛋白質的獨特的三維結構,而蛋白質結構則與它們的功能緊密相連。一些蛋白質是簡單的結構分子,如形成纖維的膠原蛋白。蛋白質可以與其他蛋白質或小分子結合;例如,作為酶的蛋白質通過與受質分子結合來執行催化其化學反應的功能。蛋白質結構是動態的;例如,血紅素在哺乳動物血液中捕捉、運輸和釋放氧氣分子的過程中能夠發生微小的結構變化。
基因序列上的單個核苷酸變化(密碼子改變)可能會導致所編碼蛋白質的胺酸序列相應改變。由於蛋白質結構是由其胺酸序列所決定的,一個胺酸的變化就有可能通過使結構失去穩定性或改變蛋白質表面而影響與該蛋白質其他蛋白質和分子的相互作用,而引起蛋白質性質發生劇烈的改變。例如,鐮刀型細胞貧血症是一種人類遺傳性疾病,是由編碼血紅素中的β-球蛋白亞基的基因中的一個核苷酸突變所引起的,這一突變導致一個胺酸發生改變從而改變了血紅素的物理性質;在這一疾病中,突變的血紅素互相結合在一起,堆積而形成纖維,從而扭曲了攜帶血紅素的紅血球的形狀。這些扭曲的鐮刀狀細胞無法在血管中通暢地流動,容易堆積而阻塞血管或者被降解,從而引起貧血疾病。
也有一些基因被轉錄為RNA分子後卻不被轉譯成蛋白質,這些RNA分子就被稱為非編碼RNA。在一些例子中,這些非編碼RNA分子(如核糖體RNA和轉運RNA)摺疊形成結構並參與部分關鍵性細胞功能。還有的RNA(如microRNA)還能夠通過與其他RNA分子雜交結合而發揮調控作用。

暹羅貓具有溫度敏感型突變,從而引起毛色的不同。
先天與後天
雖然基因含有生物體所需功能的所有資訊,環境依然在確定生物體最終的表現型中發揮著重要作用;這種兩面性被稱為「先天與後天」,也可以說,一個生物體的表現型依賴於遺傳與環境的相互作用。這種相互作用的一個例子就是溫度敏感型突變:蛋白質序列中的單個胺酸突變通常不會改變該蛋白質的行為和與其他分子的相互作用關係,但卻能夠使該蛋白質結構變得不穩定。在一個高溫環境中,分子的運動加快,分子間的碰撞也加強,這就使得這一蛋白質的結構被破壞從而無法發揮它的功能;而在一個低溫環境中,蛋白質結構卻可以保持穩定並能夠發揮正常的功能。這類突變所引起的改變在暹羅貓毛色的變化中可以被觀察到。這種貓體內一種負責生產色素的酶含有一個突變,這個突變能夠導致這種酶在高溫時變得不穩定並失去其功能。因此,在貓皮膚溫度較低處(如四肢、尾部、面部等)的毛色為深色,而較高處為淺色。
在人類遺傳疾病苯丙酮尿症中,環境因素也具有重大的影響。導致苯丙酮尿症的突變破壞了機體降解苯丙胺酸的能力,導致具有毒性的中間產物分子在體內堆積,從而引起嚴重的進行性智能發育不全和癲癇。帶有苯丙酮尿症突變的病人需要遵守嚴格的飲食,以避免攝入含苯丙胺酸的食品,才能保持正常的和健康的生活。

轉錄因子與DNA結合,影響了所結合基因的轉錄。
基因調控
一個生物體的基因組含有數千個基因,但並不是所有的基因都需要保持活化狀態。基因的表現表現為被轉錄為mRNA,然後再被轉譯成蛋白質;而細胞中存在許多方式可以來控制基因的表現,以便使蛋白質的產生符合細胞的需求。而控制基因表現「開關」的主要調控因子之一就是轉錄因子;它們是一類結合在基因的起始位點上的調控蛋白,可以活化或抑制基因的轉錄。
例如,在大腸桿菌細菌基因組內存在著一系列合成色胺酸所需的基因。然而,當細菌細胞可以從環境中獲得色胺酸時,這些基因就不被細胞所需要。色胺酸的存在直接影響了這些基因的活性,這是因為色胺酸分子會與色胺酸操縱子(一種轉錄因子)結合,引起操縱子結構變化,使得操縱子能夠結合到合成色胺酸所需基因上。色胺酸操縱子阻斷了這些基因的轉錄和表現,因而對色胺酸的合成進程產生了負反饋調控作用。
多細胞生物中的基因表現的差異性非常明顯:雖然各類細胞都含有相同的基因組,卻由於不同的基因表現而具有不同的結構和行為。多細胞生物中的所有細胞都來源於一個單一細胞,通過響應外部或細胞之間的信號而不斷分化並逐漸建立不同的基因表現規律來產生不同的行為。因為沒有一個單一基因能夠負責多細胞生物中的各個組織的發育,因此這些規律應來自於許多細胞之間的複雜的相互作用。這些過程都要通過基因調控來完成。 真核生物體內的染色質中存在著能影響基因轉錄的結構特點,常常表現為DNA和染色質的修飾形式(如DNA的甲基化),而且能夠穩定遺傳給子細胞。
這些特點是「附加性」的,因為它們存在於DNA序列的「頂端」並且可以從一個細胞遺傳給它的下一代。由於這些附加性特點,在相同培養基中生長的不同的細胞類型依然保持其不同的特性。雖然附加性特點在整個發育過程中通常是動態的,但是有一些,例如副突變(paramutation)現象可以被多代遺傳,也是DNA是遺傳的分子基礎這一通用法則的極少數例外。
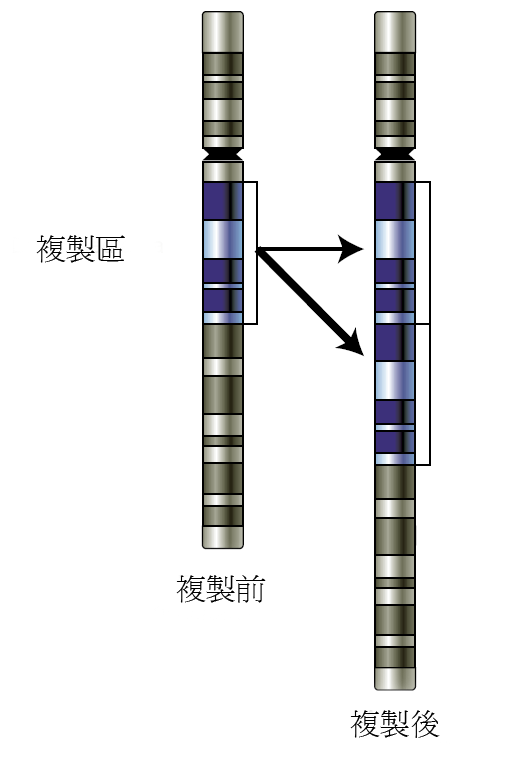
通過提供冗餘,基因複製允許發生變異:即使一個基因在複製過程中發生突變而失去原先的功能也不會對生物體造成傷害。
遺傳變化
突變
在DNA複製的過程中,第二鏈的聚合中偶爾會產生複製錯誤。這些錯誤被稱為突變,它們能夠對於一個生物體的表現型產生影響,特別是當它們位於一個基因的蛋白質編碼區中時。錯誤率通常非常低:每0.1 - 1億個鹼基才會出現1個錯誤;這是由於DNA聚合酶具有「校對」能力。
沒有校對機制則錯誤率會增加1000倍,例如許多病毒所依賴的DNA或RNA聚合酶缺乏校對能力,這使得病毒複製過程具有很高的突變率。能夠增加DNA發生改變的機率的因素被稱為突變原:一些化學品常常可以通過影響正常的鹼基對結構來提高DNA複製中的錯誤率,而紫外線能通過破壞DNA結構來誘發突變。由於對DNA的傷害在自然界中隨時都會發生,細胞則利用DNA修復機制來修復DNA中存在的錯誤配對和斷裂,但有時也無法將受破壞的DNA還原到破壞前的序列。
在利用染色體互換來交換DNA和重組基因的生物體中,減數分裂過程中所出現的配對錯誤也會導致突變。當相似序列導致姐妹染色體產生錯誤配對時,這種染色體互換出現錯誤的可能性非常大;這使得基因組中一些區域更趨向於以這一方式發生突變。這些錯誤能夠對DNA序列產生很大的結構變化:整個區域的重複、倒位或刪除,或者不同染色體之間發生意外性的交換(被稱為染色體易位)。

基於對多個同源基因序列的比較結果所構建的進化樹。
自然選擇與進化
突變會使生物體具有不同的基因型,並可能導致不同的表現型。許多突變對於生物體的表現型、健康和繁殖適應性基本沒有影響。有影響的突變則往往是有害的,但也有少量是有益的。在對果蠅的研究中發現,如果一個突變改變了基因所編碼的蛋白質,那這一突變很可能是有害的(大約有70%的此類突變具有破壞性影響,而剩餘的突變則是中性的或微弱有益的)。
群體遺傳學是研究基因變異在生物群體中的分布和這一分布的時間變化性。一個群體中的一個等位基因的變化頻率會受到自然選擇的影響,具有更高的存活率和繁殖率的等位基因能夠隨著時間而越來越頻繁地出現在該群體中。此外,遺傳漂變能夠引發等位基因出現頻率的隨機變化而不受自然選擇的影響。
在經過多個世代的傳承後,生物體的基因組會發生改變,引起被稱為進化的現象。突變和對於有益突變的選擇使得一個物種不斷地進化到能夠更好地在所處的環境中生存下來的形式,這一過程被稱為適者生存。新的物種的形成常常是由於地理分離而造成的,地理上的分離使得不同種群能夠在遺傳學上獨立發展而產生分化。遺傳學定律在群體生物學和進化研究中的應用被總結為現代綜合理論。
由於進化過程中的序列分化和變化,物種的DNA序列之間的差異可以用作「分子時鐘」(molecular clock)來計算物種之間的進化距離。遺傳比較被普遍認為是鑑定物種之間親緣關係的最準確的方法,過去常用的方法則是比較物種之間的表現型的特徵。物種之間的進化距離可以用進化樹來綜合表示,進化樹可以表示由共同祖先隨時間分化而來的物種之間的親緣關係,但不能表示無親緣關係的物種之間的遺傳物質的轉移(被稱為基因水平轉移,在細菌中非常普遍)。

黑腹果蠅(Drosophila melanogaster)是一種流行於遺傳學研究中的模式生物。
研究與技術
模式生物與遺傳學
一開始遺傳學家們的研究對象很廣泛,但逐漸地集中到一些特定物種(模式生物)的遺傳學上。這是由於新的研究者更趨向於選擇一些已經獲得廣泛研究的生物體作為研究目標,使得模式生物成為多數遺傳學研究的基礎。
模式生物的遺傳學研究包括基因調控以及發育和癌症相關基因的研究。 模式生物具有傳代時間短、易於基因操縱等優點,使得它們成為流行的遺傳學研究工具。目前廣泛使用的模式生物包括:大腸桿菌(Escherichia coli)、釀酒酵母(Saccharomyces cerevisiae)、阿拉伯芥(Arabidopsis thaliana)、線蟲(Caenorhabditis elegans)、果蠅(Drosophila melanogaster)以及小鼠(Mus musculus)。
醫學相關的遺傳學研究
醫學遺傳學的目的是了解基因變異與人類健康和疾病的關係。當尋找一個可能與某種疾病相關的未知基因時,研究者通常會用遺傳連鎖和遺傳系譜來定位基因組上與該疾病相關的區域。在群體水平上,研究者會採用孟德爾隨機法來尋找基因組上與該疾病相關的區域,這一方法也特別適用於不能被單個基因所定義的多基因性狀。一旦候選基因被發現,就需要對模式生物中的對應基因(直系同源基因)進行更多的研究。對於遺傳疾病的研究,越來越多發展起來的研究基因型的技術也被引入到藥物遺傳學中,來研究基因型如何影響藥物反應。
癌症雖然不是傳統意義上的遺傳病,但被認為是一種遺傳性疾病。癌症在機體內的產生過程是一個綜合性事件。機體內的細胞在分裂過程中有一定機率會發生突變。這些突變雖然不會遺傳給下一代,但會影響細胞的行為,在一些情況下會導致細胞更頻繁地分裂。有許多生物學機制能夠阻止這種情況的發生:信號被傳遞給這些不正常分裂的細胞並引發其凋亡;但有時更多的突變使得細胞忽略這些信號。這時機體內的自然選擇和逐漸積累起來的突變使得這些細胞開始無限制生長,從而成為癌症性腫瘤(惡性腫瘤),並侵染機體的各個器官。
相關研究技術
可以在實驗室中對DNA進行操縱。限制性內切酶是一種常用的剪切特異性序列的酶,用於製造預定的DNA片斷。然後利用DNA連接酶將這些片斷重新連接,通過將不同來源地DNA片斷連接到一起,就可以獲得重組DNA。重組DNA技術通常被用於在質體(一種短的環形DNA片斷,含有少量基因)中,這常常與基因轉殖生物的製造有關。將質體轉入細菌中,再在洋菜平板培養基上生長這些細菌(來分離菌落複製),然後研究者們就可以用複製菌落來擴增插入的質體DNA片斷(這一過程被稱為分子複製)。
DNA還能夠通過一個被稱為聚合酶鏈鎖反應(又被稱為PCR)的技術來進行擴增。利用特定的短的DNA序列,PCR技術可以分離和擴增DNA上的靶區域。因為只需要極少量的DNA就可以進行擴增,該技術也常常被用於DNA檢測(檢測特定DNA序列的存在與否)。
DNA測序與基因組
DNA測序技術是遺傳學研究中發展起來的一個最基本的技術,它使得研究者可以確定DNA片段的核苷酸序列。由弗雷德里克·桑格和他的同事於1977年發展出來的鏈終止測序法現在已經是DNA測序的常規手段。
在這一技術的幫助下,研究者們能夠對與人類疾病相關的DNA序列進行研究。 由於測序已經變得相對廉價,而且在計算機技術的輔助下,可以將大量不同片斷的序列資訊連接起來(這一過程被稱為「基因組組裝」),因此許多生物(包括人類)的基因組測序已經完成。這些技術也被用在測定人類基因組序列,使得人類基因組計劃得以在2003年完成。隨著新的高通量測序技術的發展,DNA測序的費用被大大降低,許多研究者希望能夠將測定一個人的基因組資訊的價格降到一千美元以內,從而使大眾測序成為可能。
大量測定的基因組序列資訊催生了一個新的研究領域——基因組學,研究者利用計算機軟體查找和研究生物的全基因組中存在的規律。基因組學也能夠被歸類為生物資訊學(利用計算的方法來分析生物學資料)下的一個領域。
合成生物技術
系統遺傳學(System Genetics)的應用 - 合成生物學(Synthetic Biology),後基因組時代,合成生物技術採用計算機輔助設計、全基因與基因組合成等技術,進行細胞的信號傳導與基因調控網絡的人工設計的基因工程技術,繼遺傳育種、細胞雜交、基因轉殖生物等生物技術之後,將成為21世紀遺傳工程發展的新趨勢,系統遺傳學揭示基因組的結構、功能與演化的生物系統進化機理,涉及醫學遺傳學、醫學心理學等基因系統調控、信號傳導網絡等研究,為腫瘤、遺傳病、精神病、衰老等疾病發生的診斷與藥物篩選、製藥產業等開拓了新的途徑。

恩斯特·海克爾(Ernst Haeckel)的“生命之樹”,達爾文對宇宙普遍血統的隱喻描述,是由他在德國科學界最偉大的傳播者所寫的。這是《人類進化論》(出版於1879年)中恩斯特·海克爾(Ernst Haeckel)的樹的英文版,這是海克爾(Haeckel )對生命樹的幾種描寫之一。“人”在樹冠上;對於海克爾來說,就像許多早期的進化論者一樣,人類被認為是進化的頂峰。

洋菜平板上的大腸桿菌菌落,細胞複製的一個例子,常用於分子複製。
文章定位: